

Epithelial damage and tissue γδ T cells promote a unique tumor-protective IgE response
- PMID: 30013146
- PMCID: PMC6071860
- DOI: 10.1038/s41590-018-0161-8
Epithelial damage and tissue γδ T cells promote a unique tumor-protective IgE response
Abstract
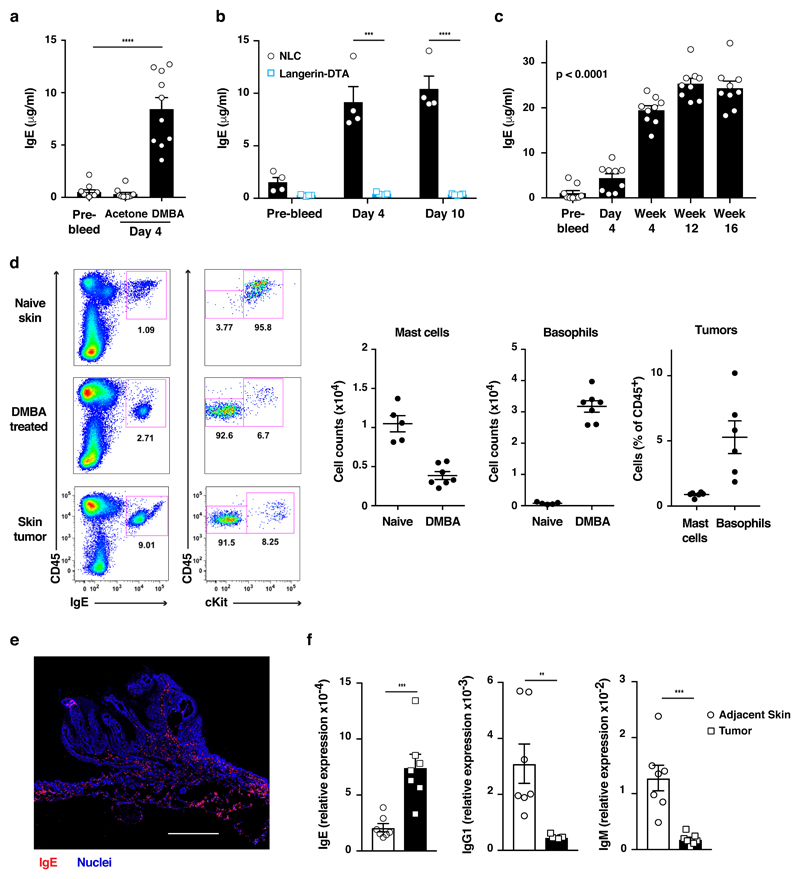
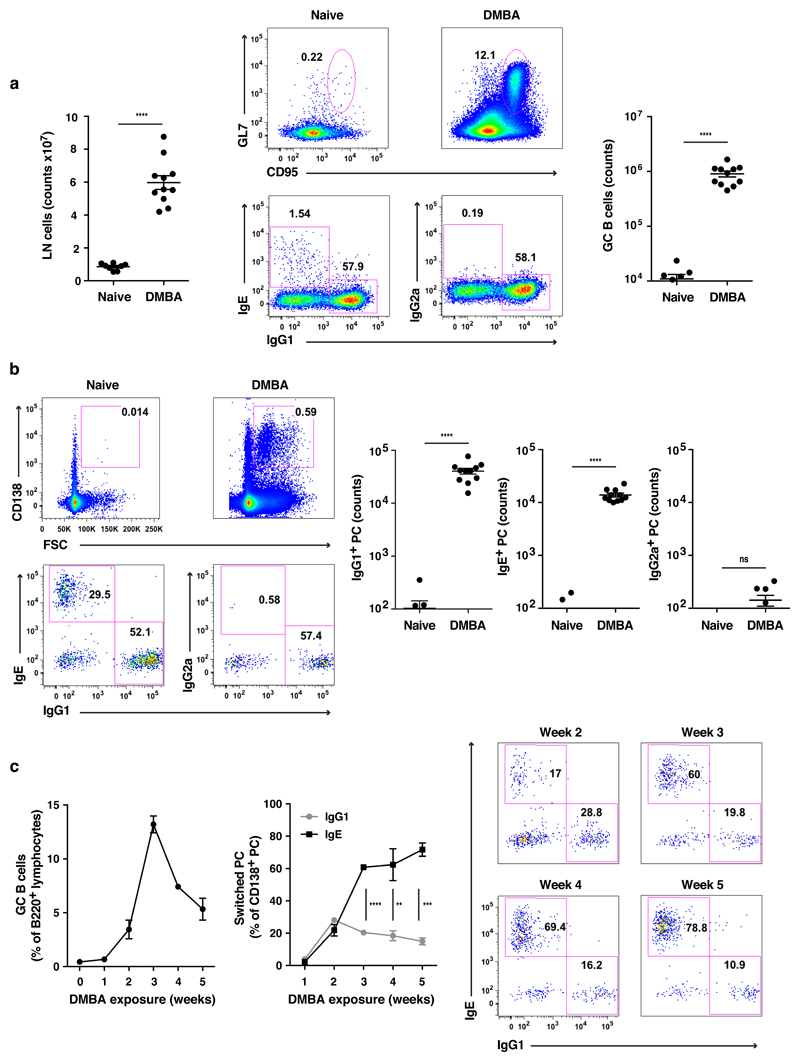
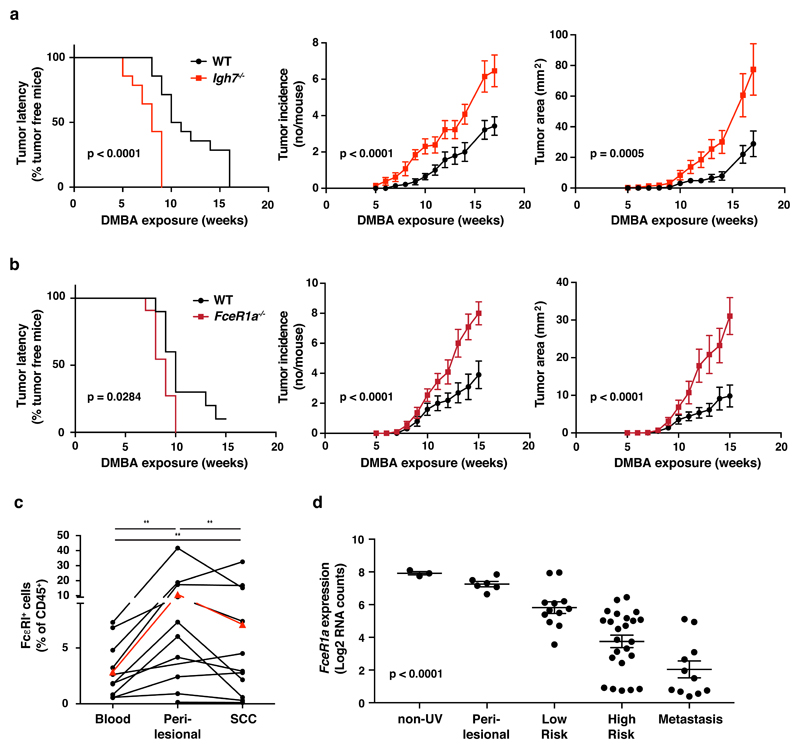
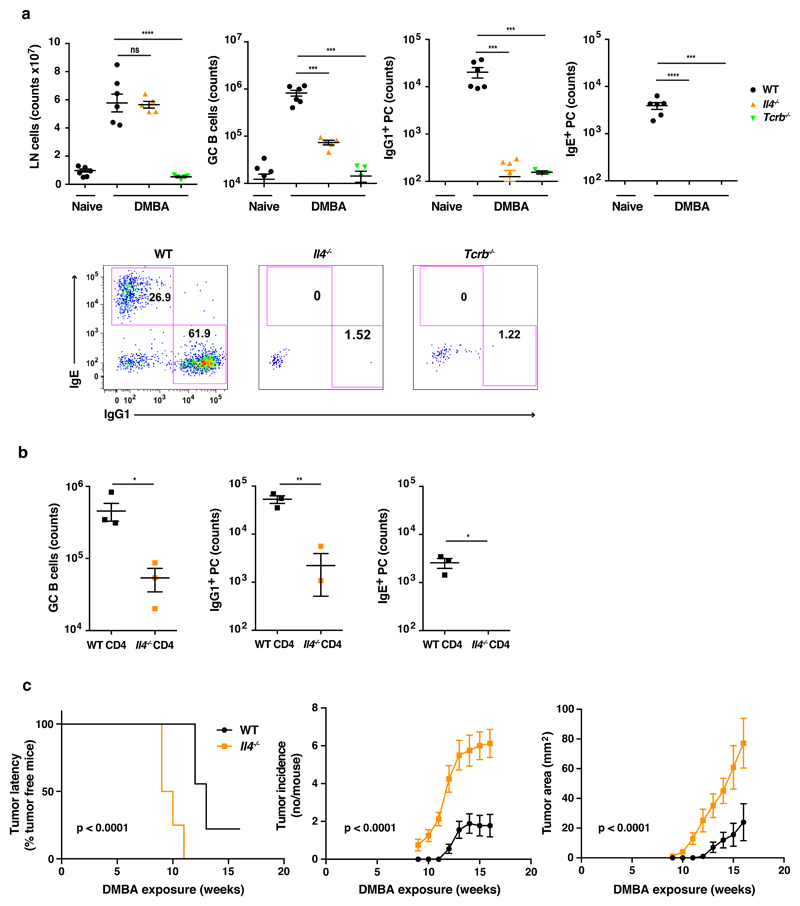
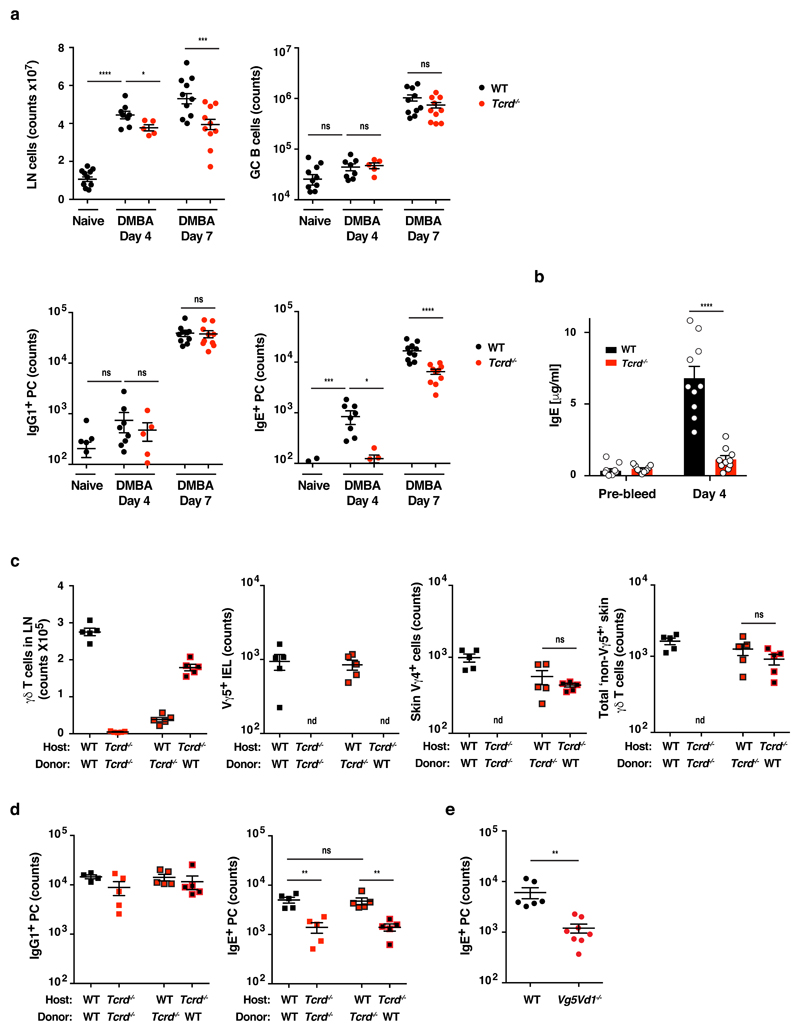
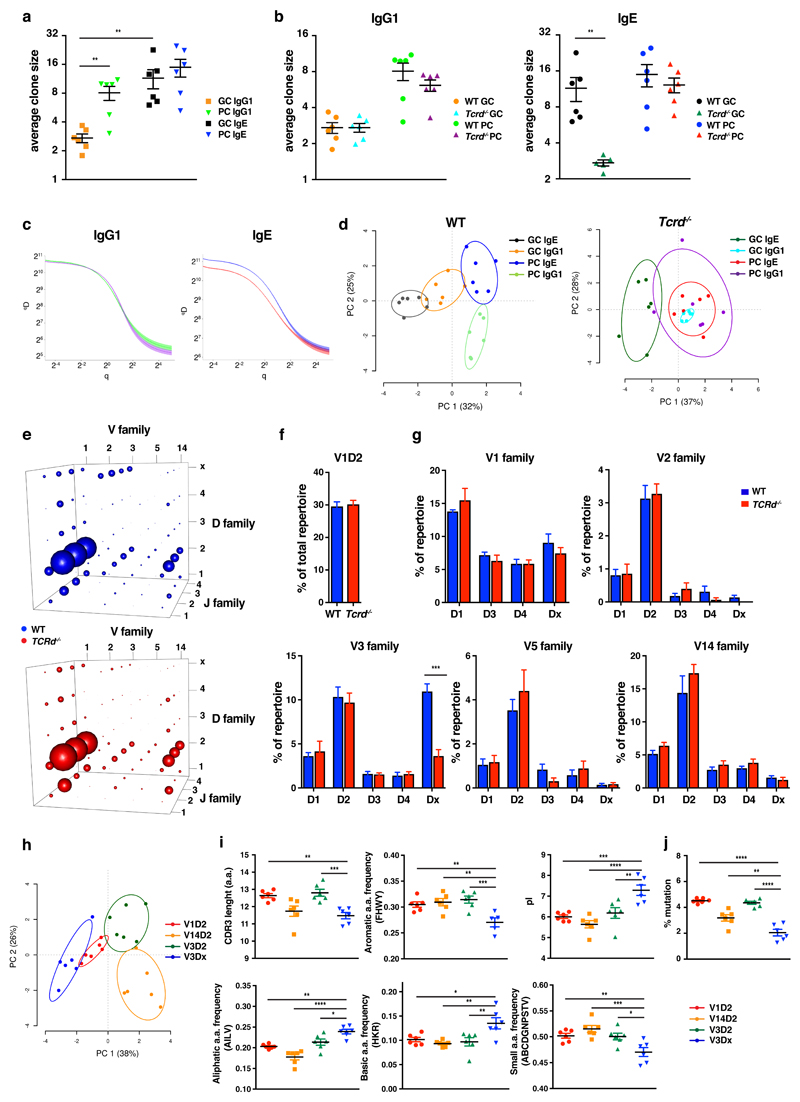
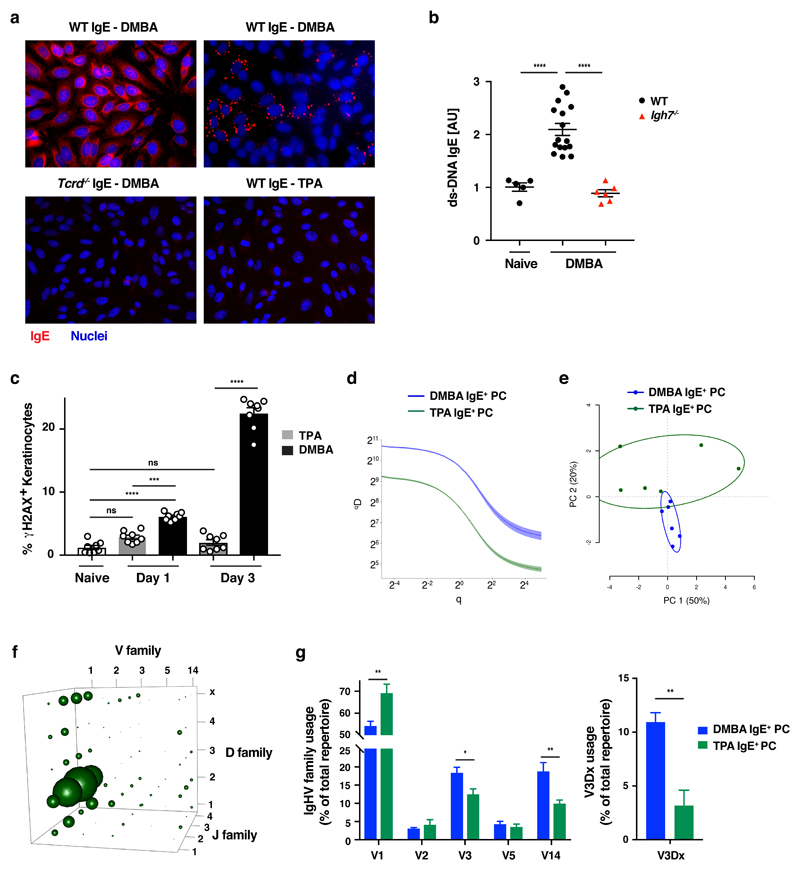
IgE is an ancient and conserved immunoglobulin isotype with potent immunological function. Nevertheless, the regulation of IgE responses remains an enigma, and evidence of a role for IgE in host defense is limited. Here we report that topical exposure to a common environmental DNA-damaging xenobiotic initiated stress surveillance by γδTCR+ intraepithelial lymphocytes that resulted in class switching to IgE in B cells and the accumulation of autoreactive IgE. High-throughput antibody sequencing revealed that γδ T cells shaped the IgE repertoire by supporting specific variable-diversity-joining (VDJ) rearrangements with unique characteristics of the complementarity-determining region CDRH3. This endogenous IgE response, via the IgE receptor FcεRI, provided protection against epithelial carcinogenesis, and expression of the gene encoding FcεRI in human squamous-cell carcinoma correlated with good disease prognosis. These data indicate a joint role for immunosurveillance by T cells and by B cells in epithelial tissues and suggest that IgE is part of the host defense against epithelial damage and tumor development.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
γδ T cells and IgE team up to prevent tumors.Nat Immunol. 2018 Aug;19(8):793-795. doi: 10.1038/s41590-018-0167-2. Nat Immunol. 2018. PMID: 30013144 No abstract available.
-
IgE in skin stress surveillance.Nat Rev Immunol. 2018 Sep;18(9):540-541. doi: 10.1038/s41577-018-0045-z. Nat Rev Immunol. 2018. PMID: 30038372 No abstract available.
References
-
- Vernersson M, Aveskogh M, Hellman L. Cloning of IgE from the echidna (Tachyglossus aculeatus) and a comparative analysis of epsilon chains from all three extant mammalian lineages. Dev Comp Immunol. 2004;28:61–75. - PubMed
-
- Profet M. The function of allergy: immunological defense against toxins. The Quarterly review of biology. 1991;66:23–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
