

Cis-regulatory determinants of MyoD function
- PMID: 30016497
- PMCID: PMC6101602
- DOI: 10.1093/nar/gky388
Cis-regulatory determinants of MyoD function
Abstract
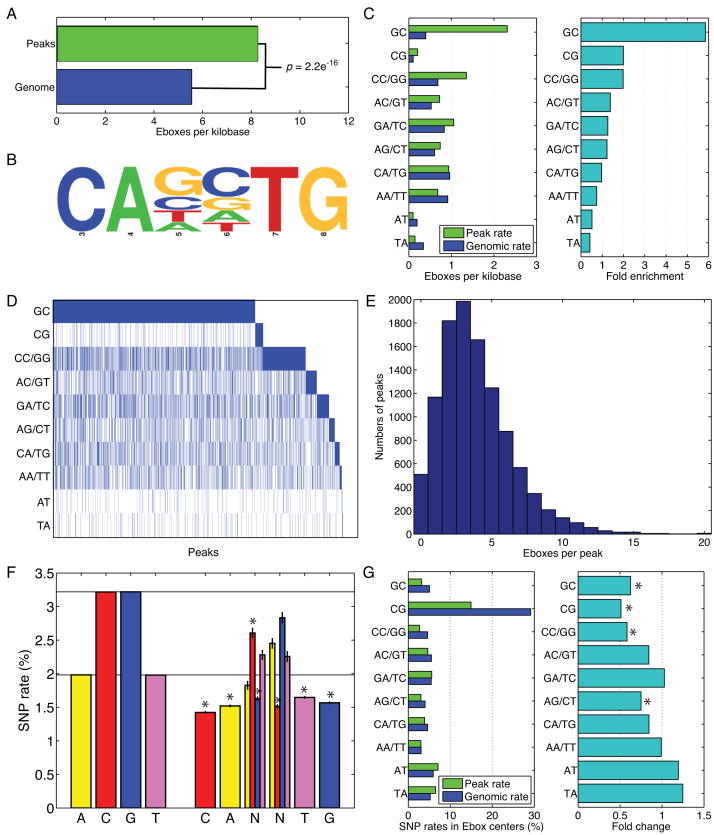
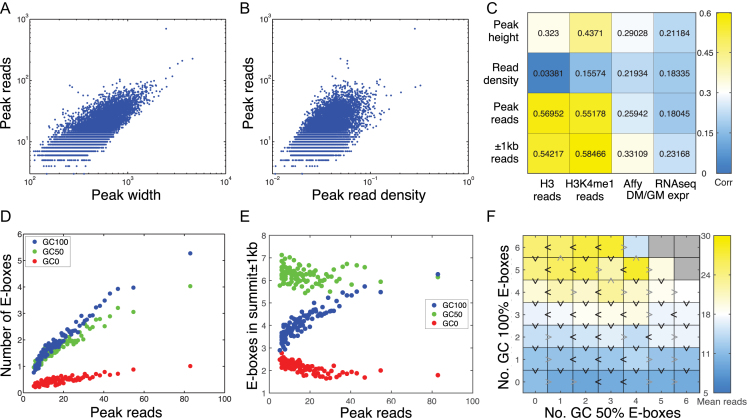
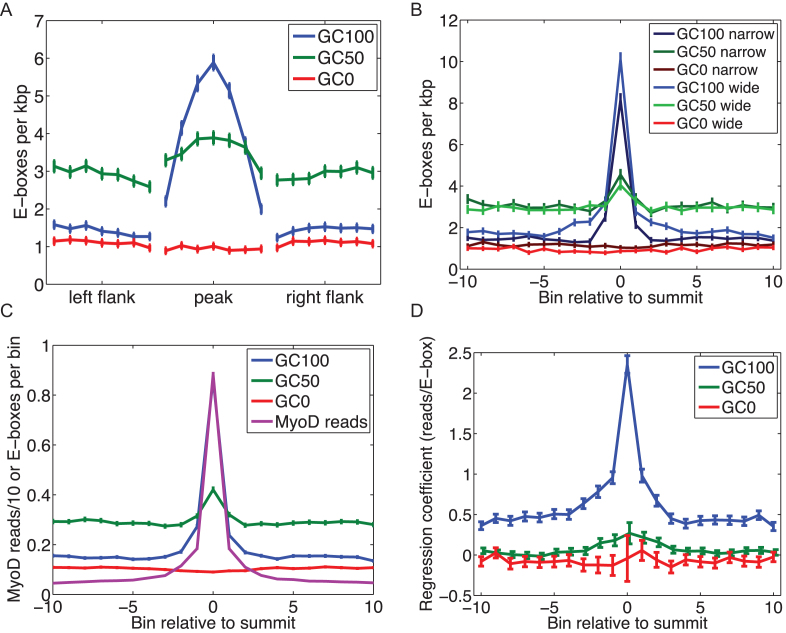
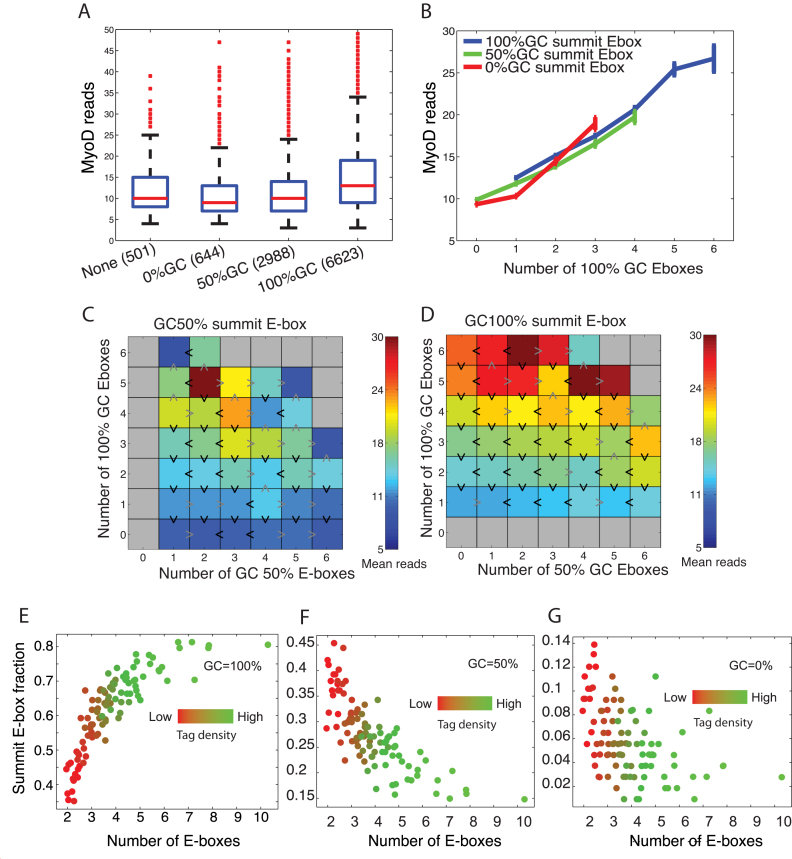
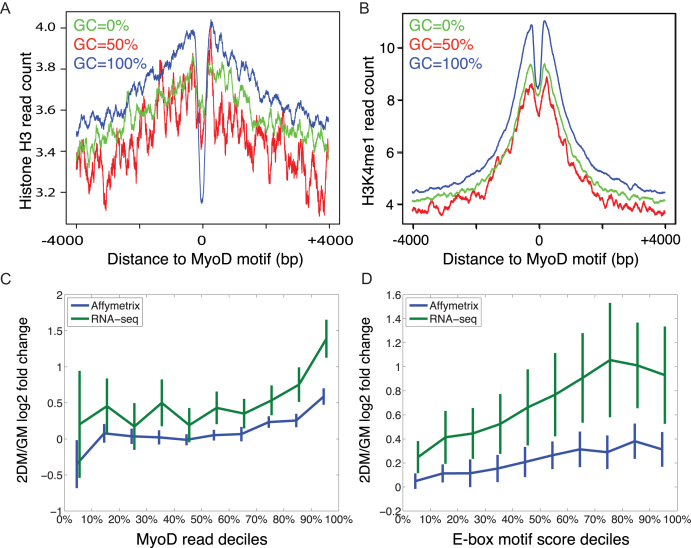
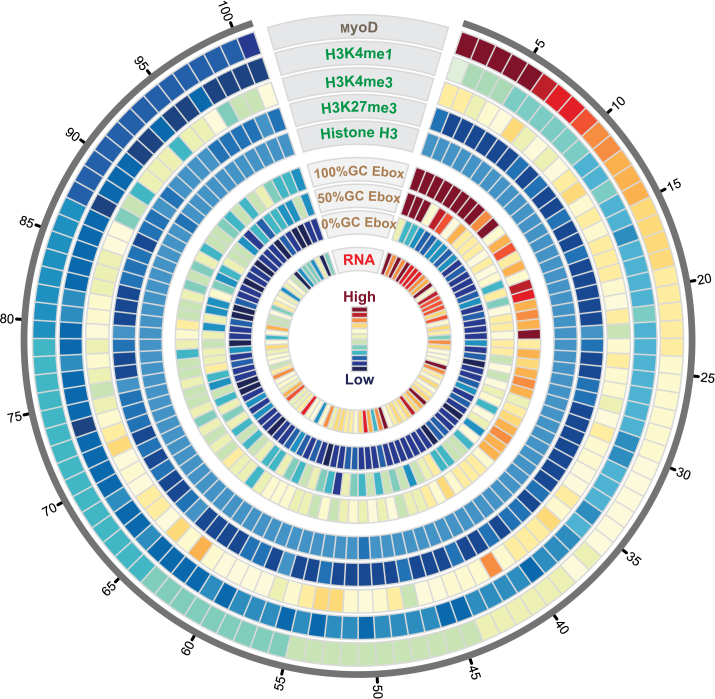
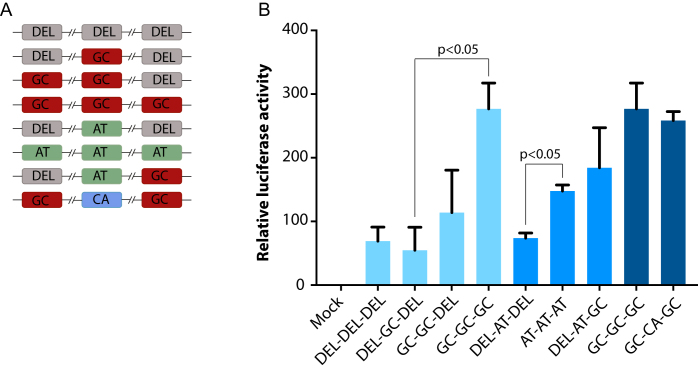
Muscle-specific transcription factor MyoD orchestrates the myogenic gene expression program by binding to short DNA motifs called E-boxes within myogenic cis-regulatory elements (CREs). Genome-wide analyses of MyoD cistrome by chromatin immnunoprecipitation sequencing shows that MyoD-bound CREs contain multiple E-boxes of various sequences. However, how E-box numbers, sequences and their spatial arrangement within CREs collectively regulate the binding affinity and transcriptional activity of MyoD remain largely unknown. Here, by an integrative analysis of MyoD cistrome combined with genome-wide analysis of key regulatory histones and gene expression data we show that the affinity landscape of MyoD is driven by multiple E-boxes, and that the overall binding affinity-and associated nucleosome positioning and epigenetic features of the CREs-crucially depend on the variant sequences and positioning of the E-boxes within the CREs. By comparative genomic analysis of single nucleotide polymorphism (SNPs) across publicly available data from 17 strains of laboratory mice, we show that variant sequences within the MyoD-bound motifs, but not their genome-wide counterparts, are under selection. At last, we show that the quantitative regulatory effect of MyoD binding on the nearby genes can, in part, be predicted by the motif composition of the CREs to which it binds. Taken together, our data suggest that motif numbers, sequences and their spatial arrangement within the myogenic CREs are important determinants of the cis-regulatory code of myogenic CREs.
Figures
References
-
- Davis R.L., Weintraub H., Lassar A.B.. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell. 1987; 51:987–1000. - PubMed
-
- Weintraub H., Genetta T., Kadesch T.. Tissue-specific gene activation by MyoD: determination of specificity by cis-acting repression elements. Genes Dev. 1994; 8:2203–2211. - PubMed
-
- Ma P.C., Rould M.A., Weintraub H., Pabo C.O.. Crystal structure of MyoD bHLH domain-DNA complex: perspectives on DNA recognition and implications for transcriptional activation. Cell. 1994; 77:451–459. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
