

Analysis of genetic polymorphisms and tropism in East African Leishmania donovani by Amplified Fragment Length Polymorphism and kDNA minicircle sequencing
- PMID: 30016714
- PMCID: PMC6218636
- DOI: 10.1016/j.meegid.2018.07.016
Analysis of genetic polymorphisms and tropism in East African Leishmania donovani by Amplified Fragment Length Polymorphism and kDNA minicircle sequencing
Abstract
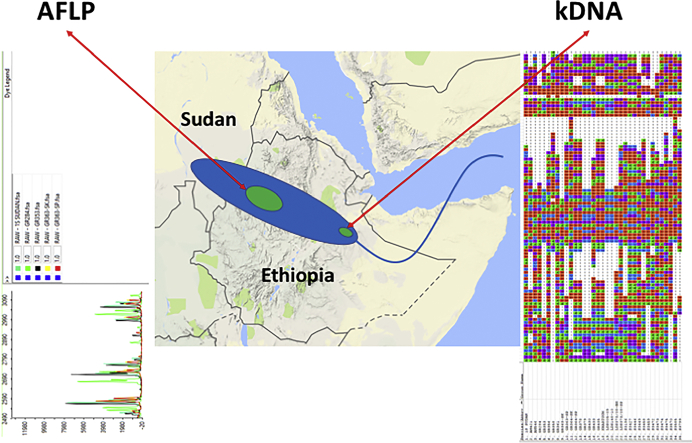
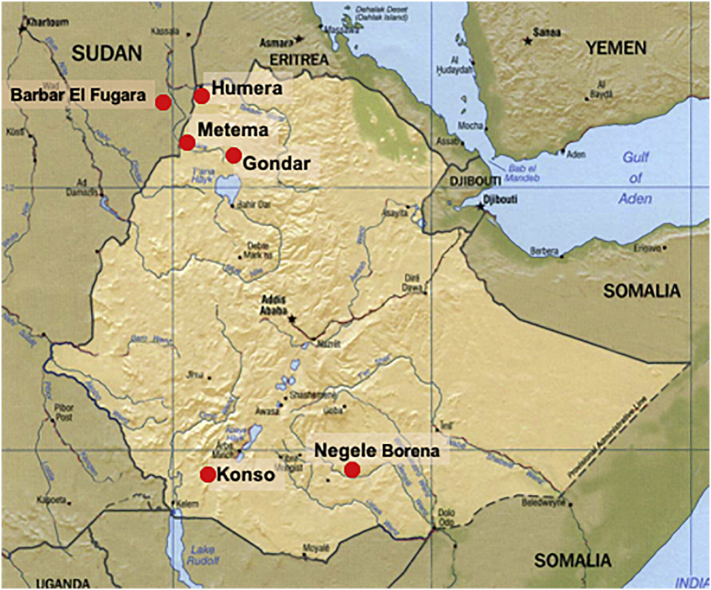
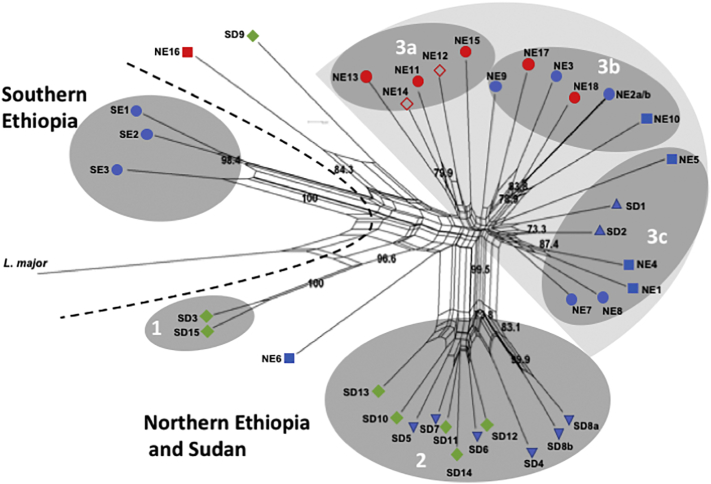
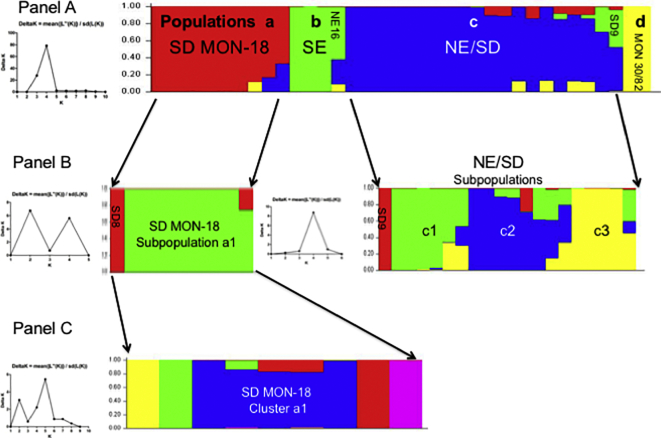
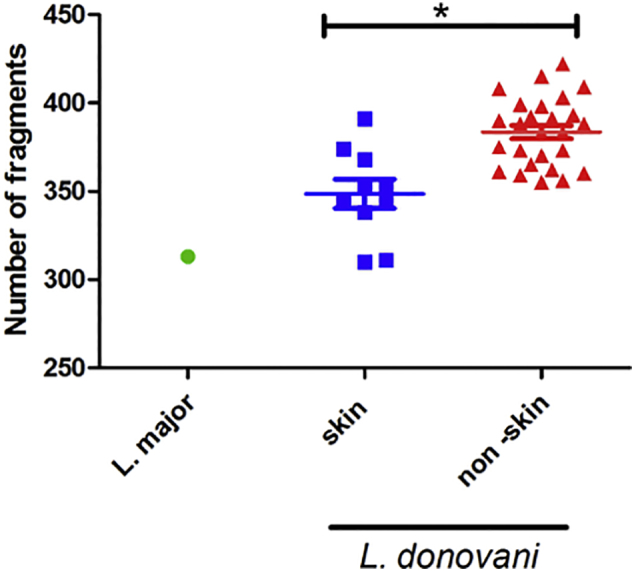
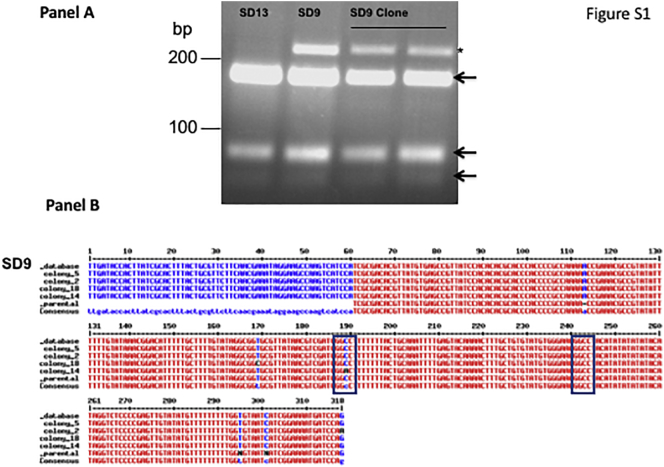
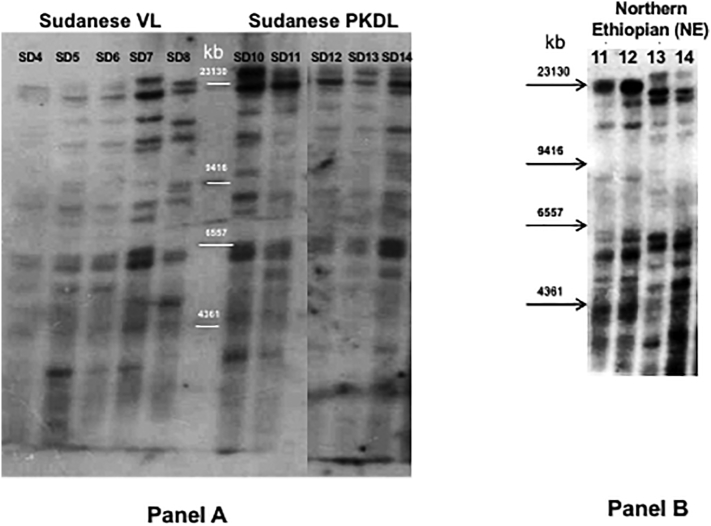
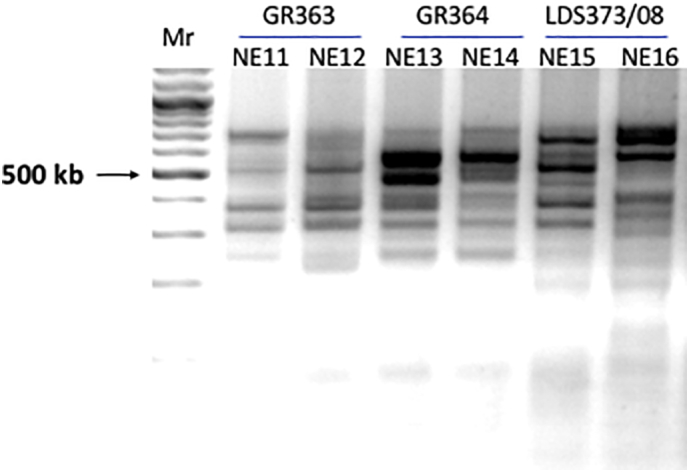
Visceral leishmaniasis (VL), the most severe form of leishmaniasis, is caused by Leishmania donovani. In addition to fatal VL, these parasites also cause skin diseases in immune-competent and -suppressed people, post-kala azar dermal leishmaniasis (PKDL) and HIV/VL co-infections, respectively. Genetic polymorphism in 36 Ethiopian and Sudanese L. donovani strains from VL, PKDL and HIV/VL patients was examined using Amplified Fragment Length Polymorphism (AFLP), kDNA minicircle sequencing and Southern blotting. Strains were isolated from different patient tissues: in VL from lymph node, spleen or bone marrow; and in HIV/VL from skin, spleen or bone marrow. When VL and PKDL strains from the same region in Sudan were examined by Southern blotting using a DNA probe to the L. donovani 28S rRNA gene only minor differences were observed. kDNA sequence analysis distributed the strains in no particular order among four clusters (A - D), while AFLP analysis grouped the strains according to geographical origin into two major clades, Southern Ethiopia (SE) and Sudan/Northern Ethiopia (SD/NE). Strains in the latter clade were further divided into subpopulations by zymodeme, geography and year of isolation, but not by clinical symptoms. However, skin isolates showed significantly (p < 0.0001) fewer polymorphic AFLP fragments (average 10 strains = 348.6 ± 8.1) than VL strains (average 26 strains = 383.5 ± 3.8).
Keywords: Amplified Fragment Length Polymorphism; Genetic polymorphism; HIV-visceral leishmaniasis co-infections; Leishmania donovani; Post-kala azar dermal leishmaniasis; kDNA minicircle sequence.
Copyright © 2018 The Authors. Published by Elsevier B.V. All rights reserved.
Figures

References
-
- Alvar J., Bashaye S., Argaw D., Cruz I., Aparicio P., Kassa A., Orfanos G., Parreno F., Babaniyi O., Gudeta N., Canavate C., Bern C. Kala-azar outbreak in Libo Kemkem, Ethiopia: epidemiologic and parasitologic assessment. Am. J. Trop. Med. Hyg. 2007;77:275–282. - PubMed
-
- Anders G., Eisenberger C.L., Jonas F., Greenblatt C.L. Distinguishing Leishmania tropica and Leishmania major in the Middle East using the polymerase chain reaction with kinetoplast DNA-specific primers. Trans. R. Soc. Trop. Med. Hyg. 2002;96:S87–S92. Suppl 1. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
