

Unique BIR domain sets determine inhibitor of apoptosis protein-driven cell death and NOD2 complex signal specificity
- PMID: 30018081
- PMCID: PMC6066185
- DOI: 10.1126/scisignal.aao3964
Unique BIR domain sets determine inhibitor of apoptosis protein-driven cell death and NOD2 complex signal specificity
Abstract
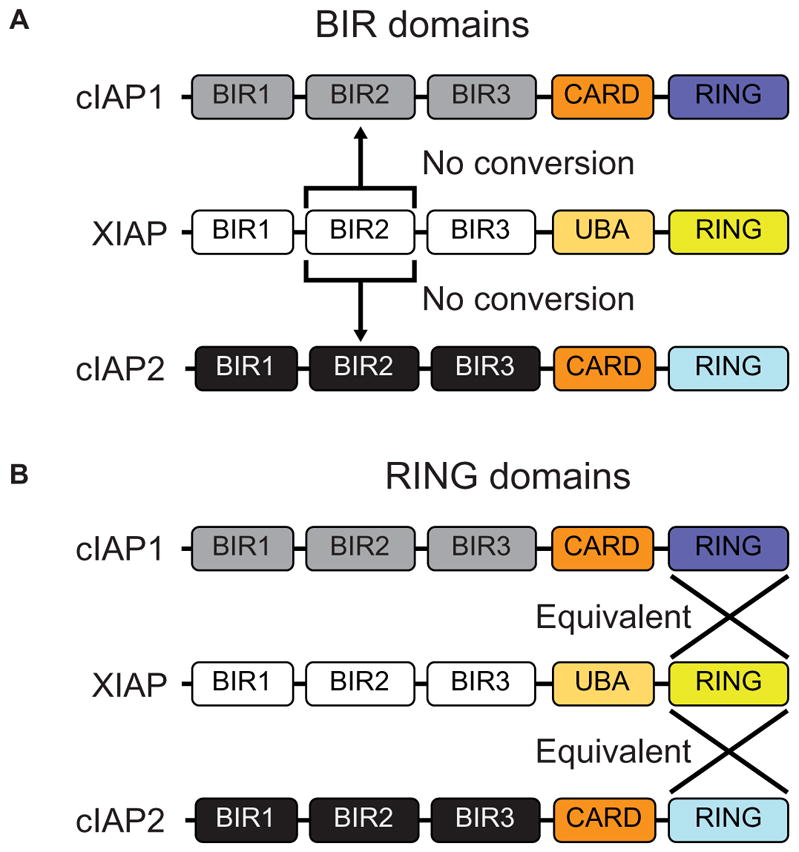
The mammalian IAPs, X-linked inhibitor of apoptosis protein (XIAP) and cellular inhibitor of apoptosis protein 1 and 2 (cIAP1 and cIAP2), play pivotal roles in innate immune signaling and inflammatory homeostasis, often working in parallel or in conjunction at a signaling complex. IAPs direct both nucleotide-binding oligomerization domain-containing 2 (NOD2) signaling complexes and cell death mechanisms to appropriately regulate inflammation. Although it is known that XIAP is critical for NOD2 signaling and that the loss of cIAP1 and cIAP2 blunts NOD2 activity, it is unclear whether these three highly related proteins can compensate for one another in NOD2 signaling or in mechanisms governing apoptosis or necroptosis. This potential redundancy is critically important, given that genetic loss of XIAP causes both very early onset inflammatory bowel disease and X-linked lymphoproliferative syndrome 2 (XLP-2) and that the overexpression of cIAP1 and cIAP2 is linked to both carcinogenesis and chemotherapeutic resistance. Given the therapeutic interest in IAP inhibition and the potential toxicities associated with disruption of inflammatory homeostasis, we used synthetic biology techniques to examine the functional redundancies of key domains in the IAPs. From this analysis, we defined the features of the IAPs that enable them to function at overlapping signaling complexes but remain independent and functionally exclusive in their roles as E3 ubiquitin ligases in innate immune and inflammatory signaling.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
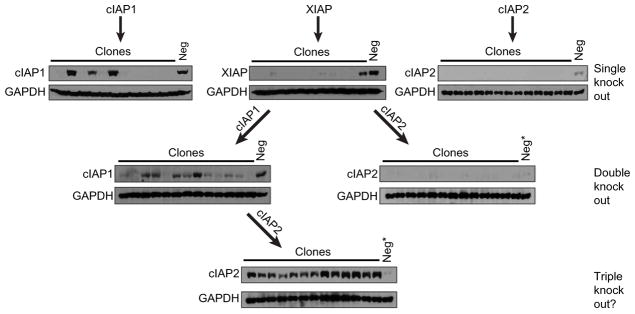
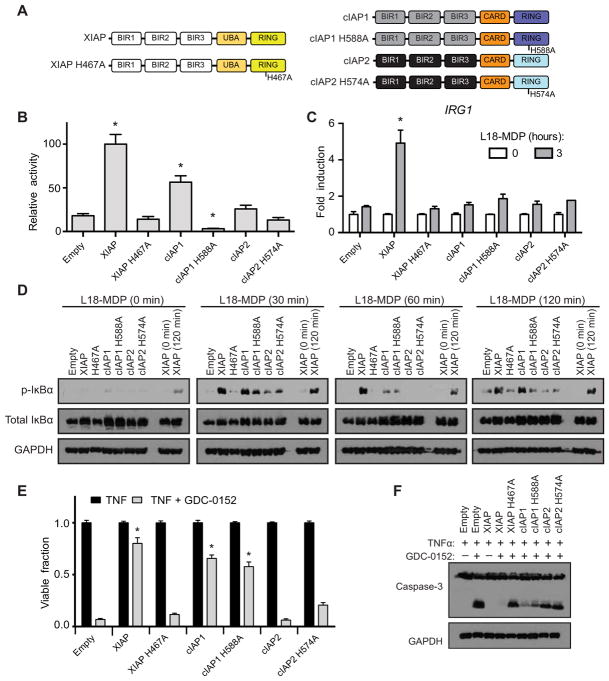
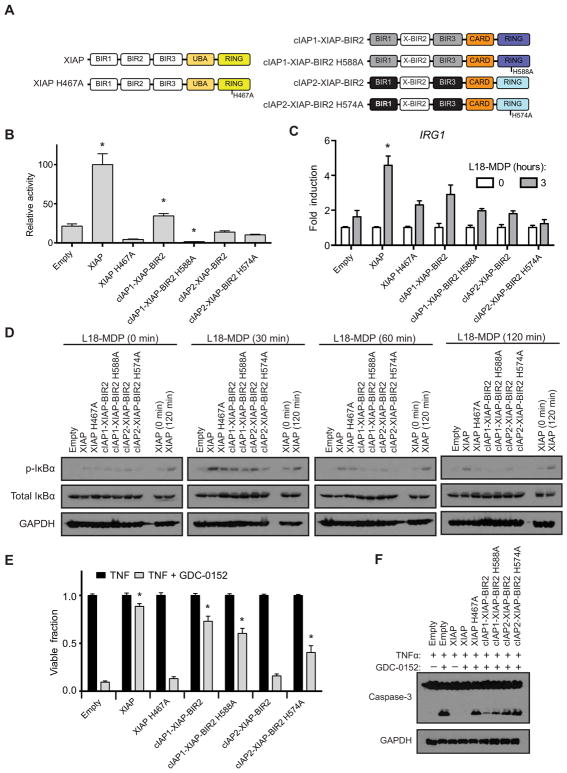
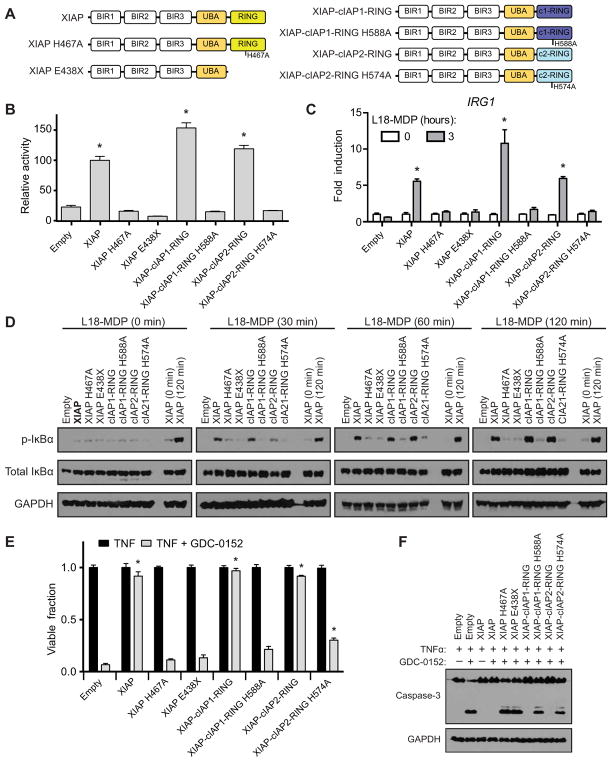
References
-
- Uren AG, Coulson EJ, Vaux DL. Conservation of baculovirus inhibitor of apoptosis repeat proteins (BIRPs) in viruses, nematodes, vertebrates and yeasts. Trends Biochem Sci. 1998;23:159–162. - PubMed
-
- Deveraux QL, Takahashi R, Salvesen GS, Reed JC. X-linked IAP is a direct inhibitor of cell-death proteases. Nature. 1997;388:300–304. - PubMed
-
- Salvesen GS, Duckett CS. IAP proteins: Blocking the road to death’s door. Nat Rev Mol Cell Biol. 2002;3:401–410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
