

Chlamydomonas Basal Bodies as Flagella Organizing Centers
- PMID: 30018231
- PMCID: PMC6070942
- DOI: 10.3390/cells7070079
Chlamydomonas Basal Bodies as Flagella Organizing Centers
Abstract
During ciliogenesis, centrioles convert to membrane-docked basal bodies, which initiate the formation of cilia/flagella and template the nine doublet microtubules of the flagellar axoneme. The discovery that many human diseases and developmental disorders result from defects in flagella has fueled a strong interest in the analysis of flagellar assembly. Here, we will review the structure, function, and development of basal bodies in the unicellular green alga Chlamydomonas reinhardtii, a widely used model for the analysis of basal bodies and flagella. Intraflagellar transport (IFT), a flagella-specific protein shuttle critical for ciliogenesis, was first described in C. reinhardtii. A focus of this review will be on the role of the basal bodies in organizing the IFT machinery.
Keywords: axoneme; bld10; bld12; bld2; central pair; centrin; centriole; intraflagellar transport (IFT); microtubules; striated fiber assemblin (SFA).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
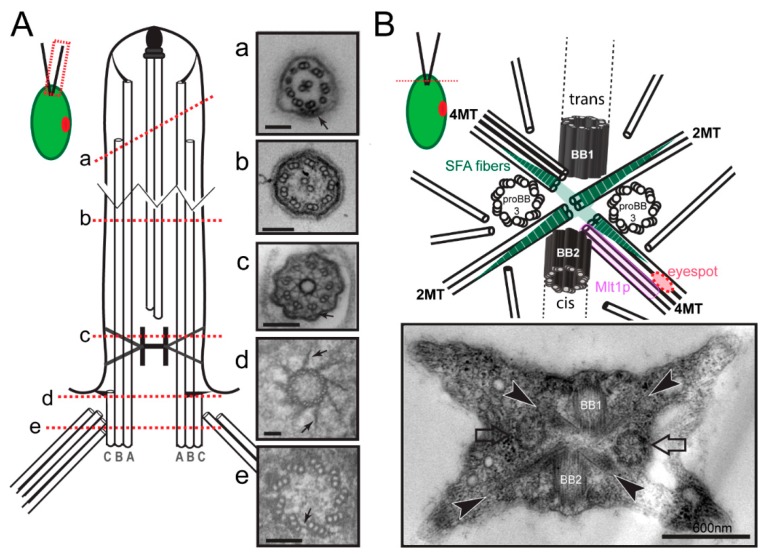
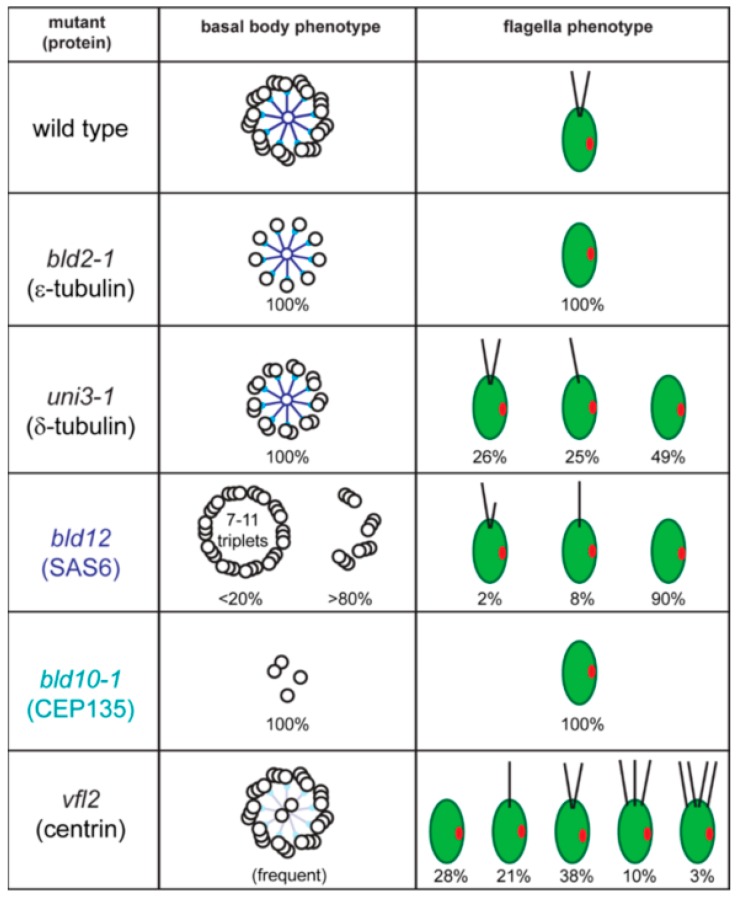
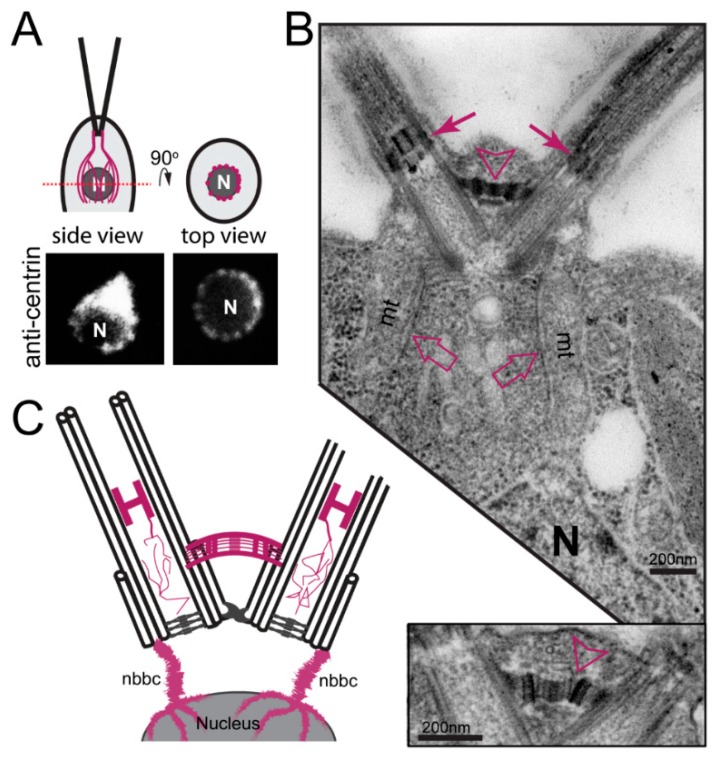
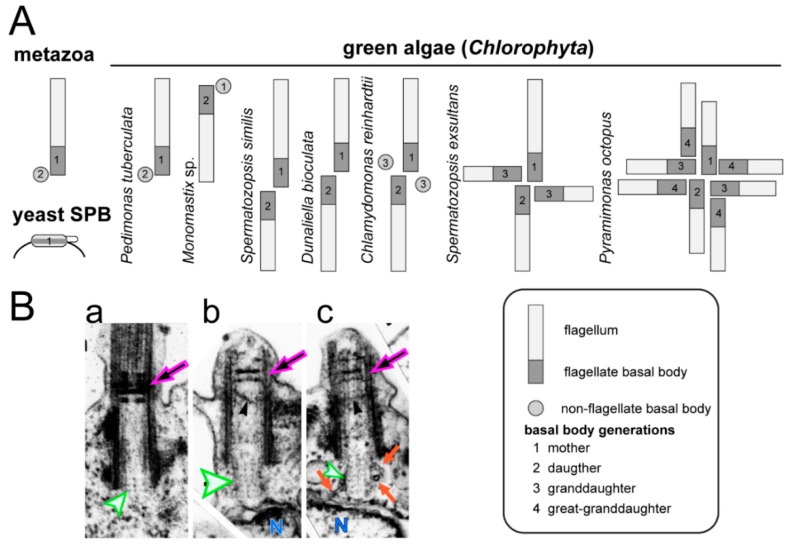
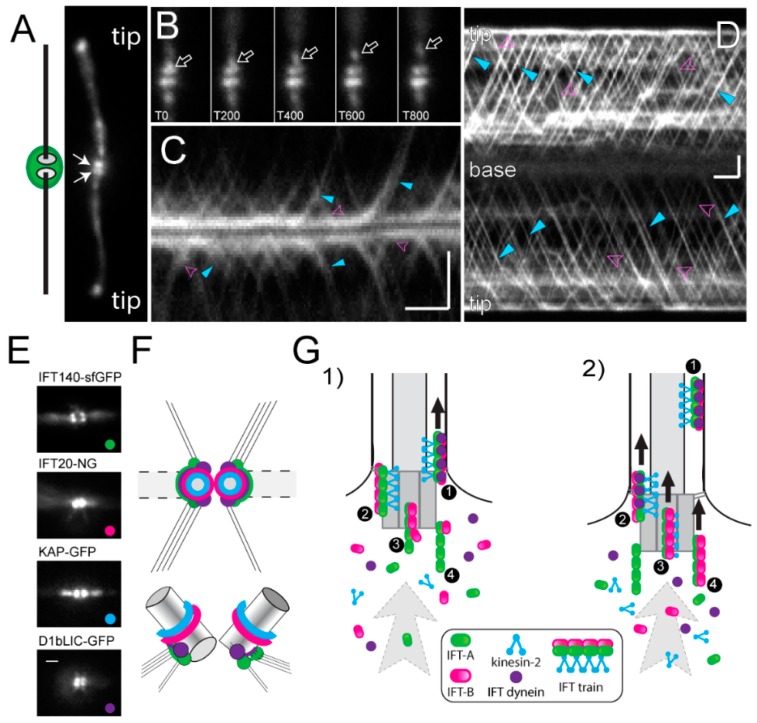
References
-
- Fawcett D.W., Porter K.R. A study of the fine structure of ciliated epithelia. J. Morphol. 1954;94:221–281. doi: 10.1002/jmor.1050940202. - DOI
-
- Musgrave A., de Wildt P., van Etten I., Pijst H., Scholma C., Kooyman R., Homan W., van den Ende H. Evidence for a functional membrane barrier in the transition zone between the flagellum and cell body of Chlamydomonas eugametos gametes. Planta. 1986;167:544–553. doi: 10.1007/BF00391231. - DOI - PubMed
-
- Dentler W.L. Structures linking the tips of ciliary and flagellar microtubules to the membrane. J. Cell Sci. 1980;42:207–220. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
