

Polyploidy in tissue homeostasis and regeneration
- PMID: 30021843
- PMCID: PMC10682953
- DOI: 10.1242/dev.156034
Polyploidy in tissue homeostasis and regeneration
Abstract
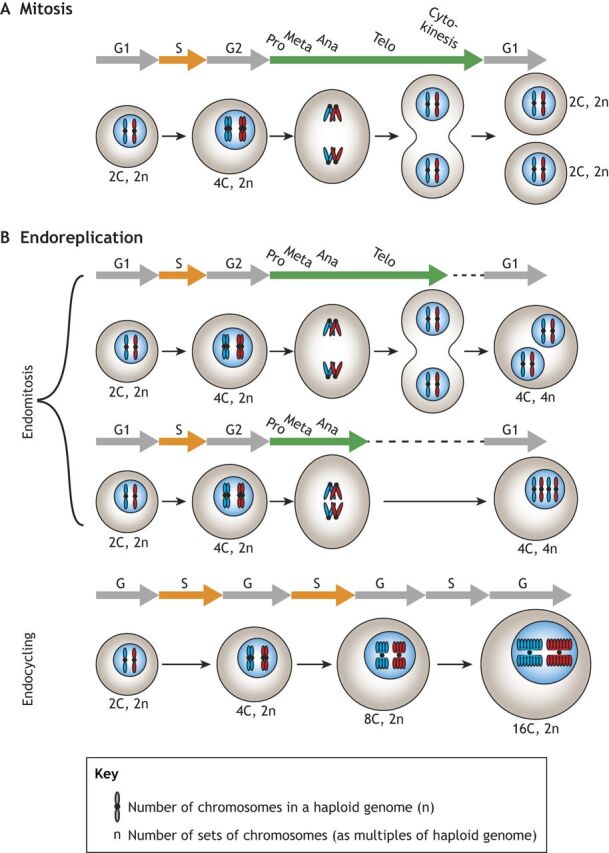
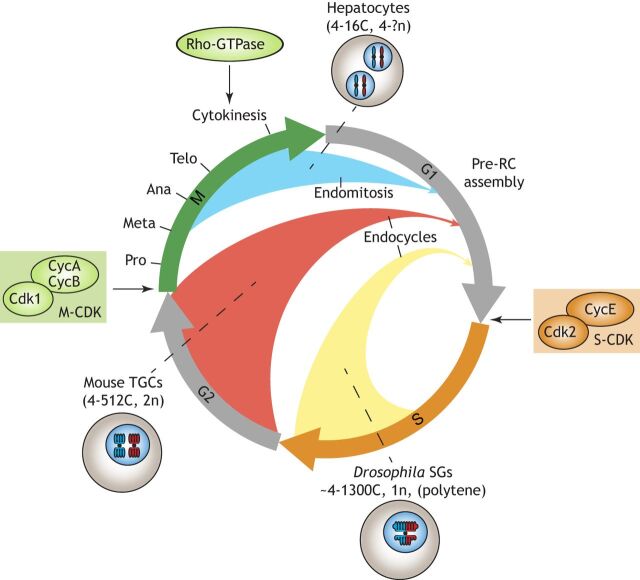
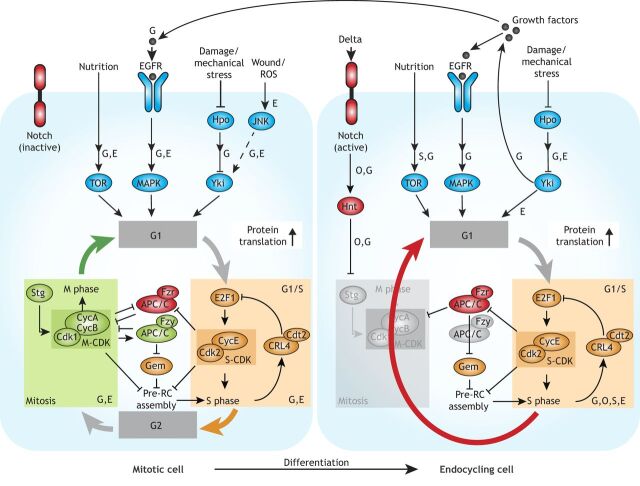
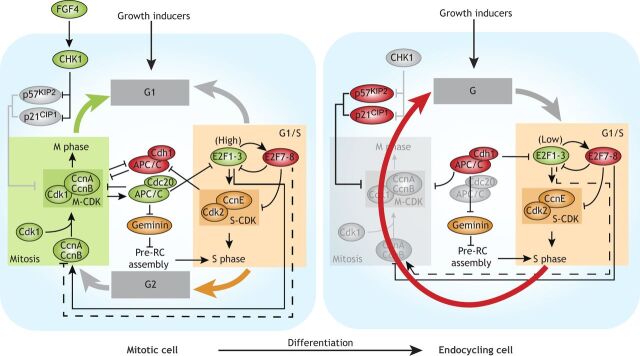
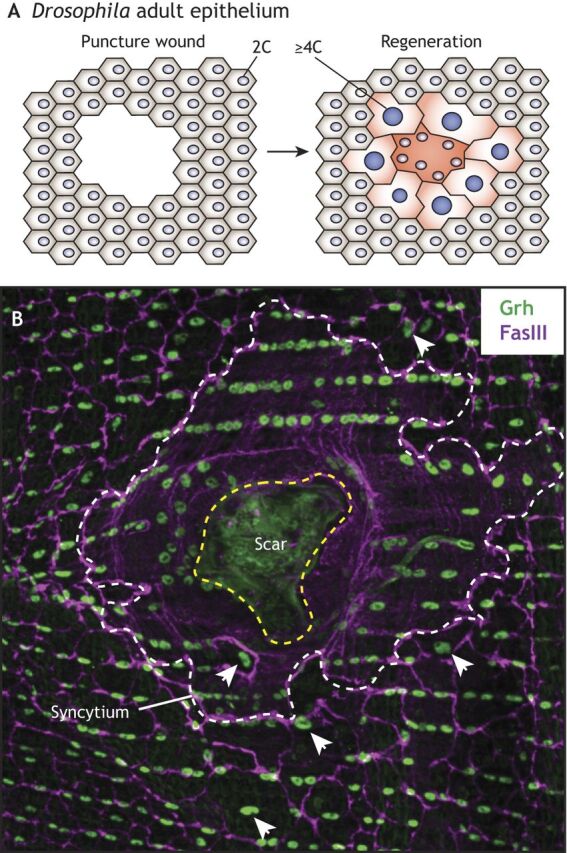
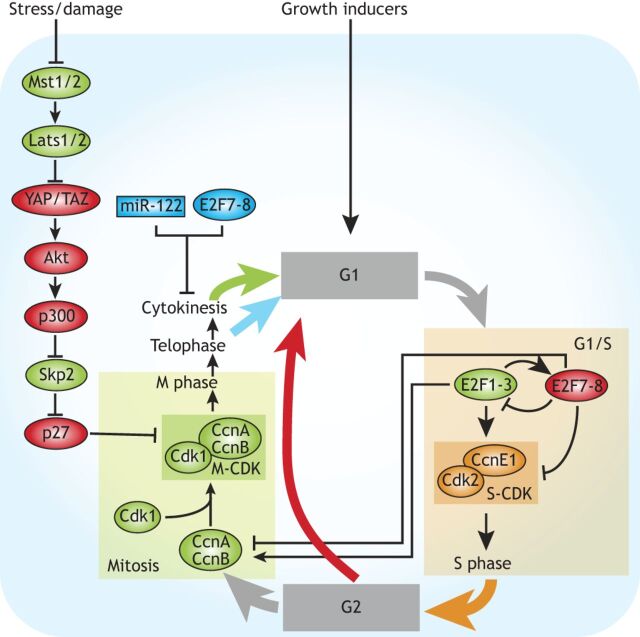
Polyploid cells, which contain multiple copies of the typically diploid genome, are widespread in plants and animals. Polyploidization can be developmentally programmed or stress induced, and arises from either cell-cell fusion or a process known as endoreplication, in which cells replicate their DNA but either fail to complete cytokinesis or to progress through M phase entirely. Polyploidization offers cells several potential fitness benefits, including the ability to increase cell size and biomass production without disrupting cell and tissue structure, and allowing improved cell longevity through higher tolerance to genomic stress and apoptotic signals. Accordingly, recent studies have uncovered crucial roles for polyploidization in compensatory cell growth during tissue regeneration in the heart, liver, epidermis and intestine. Here, we review current knowledge of the molecular pathways that generate polyploidy and discuss how polyploidization is used in tissue repair and regeneration.
Keywords: Endocycling; Endomitosis; Healing; Hippo; Polyploid; Wound.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
