

Histone Interaction Landscapes Visualized by Crosslinking Mass Spectrometry in Intact Cell Nuclei
- PMID: 30021884
- PMCID: PMC6166682
- DOI: 10.1074/mcp.RA118.000924
Histone Interaction Landscapes Visualized by Crosslinking Mass Spectrometry in Intact Cell Nuclei
Abstract
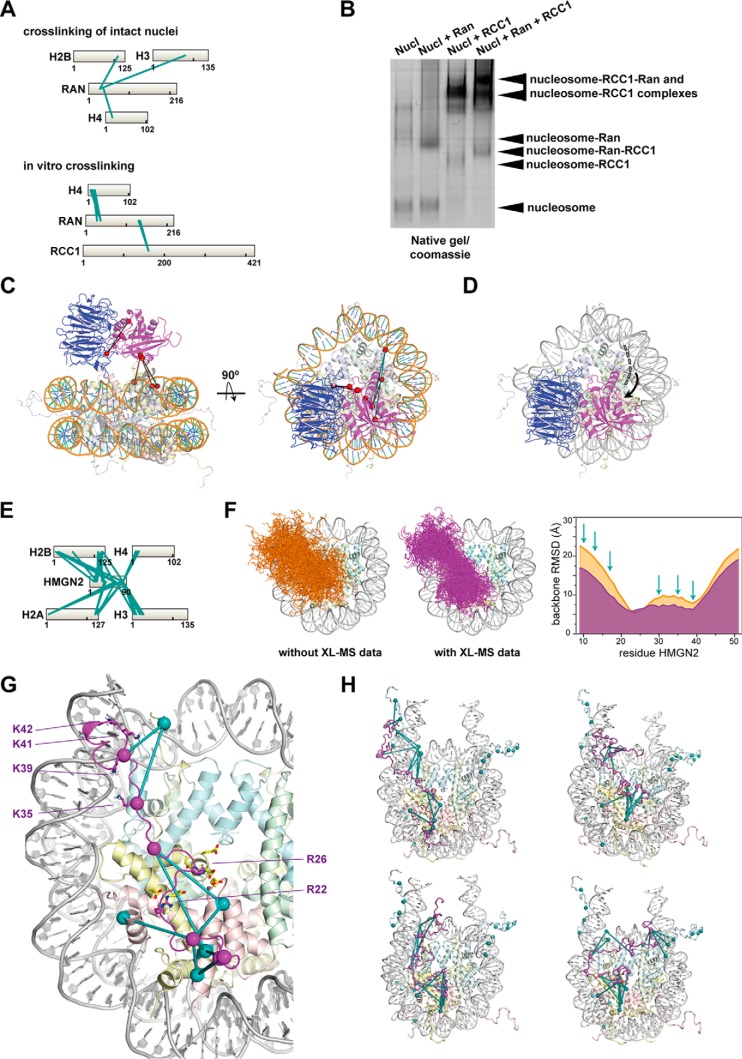
Cells organize their actions partly through tightly controlled protein-protein interactions-collectively termed the interactome. Here we use crosslinking mass spectrometry (XL-MS) to chart the protein-protein interactions in intact human nuclei. Overall, we identified ∼8,700 crosslinks, of which 2/3 represent links connecting distinct proteins. From these data, we gain insights on interactions involving histone proteins. We observed that core histones on the nucleosomes expose well-defined interaction hot spots. For several nucleosome-interacting proteins, such as USF3 and Ran GTPase, the data allowed us to build low-resolution models of their binding mode to the nucleosome. For HMGN2, the data guided the construction of a refined model of the interaction with the nucleosome, based on complementary NMR, XL-MS, and modeling. Excitingly, the analysis of crosslinks carrying posttranslational modifications allowed us to extract how specific modifications influence nucleosome interactions. Overall, our data depository will support future structural and functional analysis of cell nuclei, including the nucleoprotein assemblies they harbor.
Keywords: Chromatin function or biology; Histone interaction network – PTM interplay; Histone variant specific interactions; Histones; Mass Spectrometry; Nuclear interactome; Protein Cross-linking; Protein-Protein Interactions.
© 2018 Fasci et al.
Conflict of interest statement
Conflict of interest: The authors declare that they have no conflict of interest.
Figures





References
-
- Alberts B. (1998) The cell as a collection of protein machines: Preparing the next generation of molecular biologists. Cell 92, 291–294 - PubMed
-
- Robinson C. V., Sali A., and Baumeister W. (2007) The molecular sociology of the cell. Nature 450, 973–982 - PubMed
-
- Gavin A. C., Aloy P., Grandi P., Krause R., Boesche M., Marzioch M., Rau C., Jensen L. J., Bastuck S., Dümpelfeld B., Edelmann A., Heurtier M. A., Hoffman V., Hoefert C., Klein K., Hudak M., Michon A. M., Schelder M., Schirle M., Remor M., Rudi T., Hooper S., Bauer A., Bouwmeester T., Casari G., Drewes G., Neubauer G., Rick J. M., Kuster B., Bork P., Russell R. B., and Superti-Furga G. (2006) Proteome survey reveals modularity of the yeast cell machinery. Nature 440, 631–636 - PubMed
-
- Huttlin E. L., Ting L., Bruckner R. J., Gebreab F., Gygi M. P., Szpyt J., Tam S., Zarraga G., Colby G., Baltier K., Dong R., Guarani V., Vaites L. P., Ordureau A., Rad R., Erickson B. K., Wühr M., Chick J., Zhai B., Kolippakkam D., Mintseris J., Obar R. A., Harris T., Artavanis-Tsakonas S., Sowa M. E., De Camilli P., Paulo J. A., Harper J. W., and Gygi S. P. (2015) The BioPlex Network: A systematic exploration of the human interactome. Cell 162, 425–440 - PMC - PubMed
-
- Hein M. Y., Hubner N. C., Poser I., Cox J., Nagaraj N., Toyoda Y., Gak I. A., Weisswange I., Mansfeld J., Buchholz F., Hyman A. A., and Mann M. (2015) A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell 163, 712–723 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
