

The Neuronal Basis of Predictive Coding Along the Auditory Pathway: From the Subcortical Roots to Cortical Deviance Detection
- PMID: 30022729
- PMCID: PMC6053868
- DOI: 10.1177/2331216518784822
The Neuronal Basis of Predictive Coding Along the Auditory Pathway: From the Subcortical Roots to Cortical Deviance Detection
Abstract
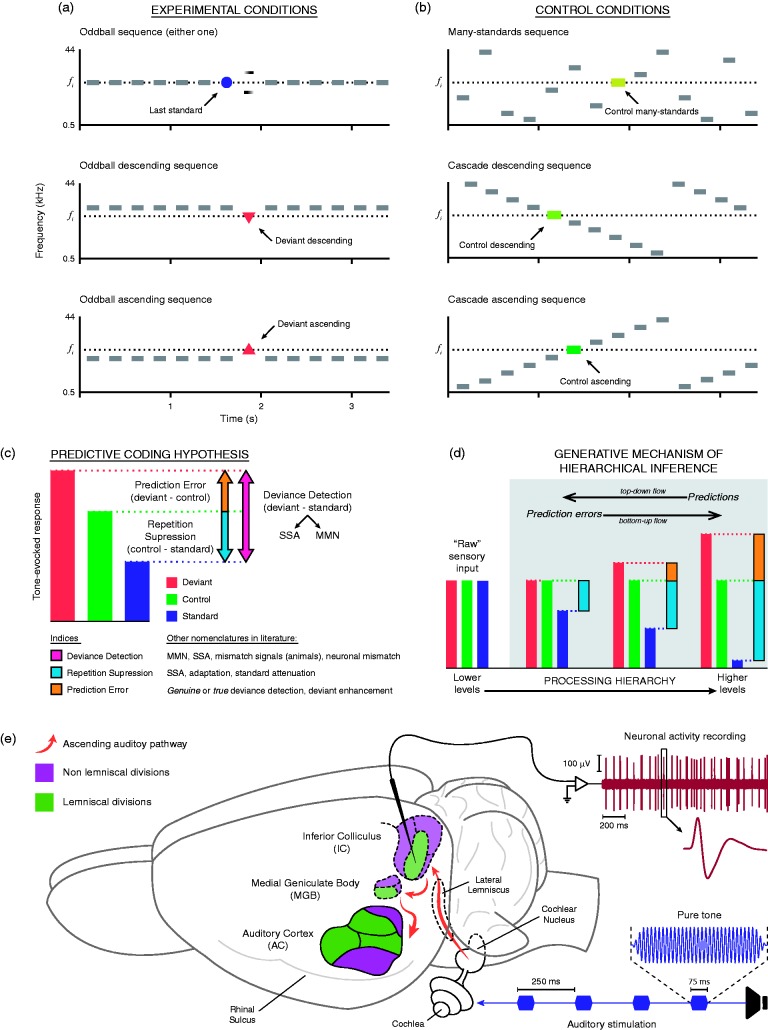
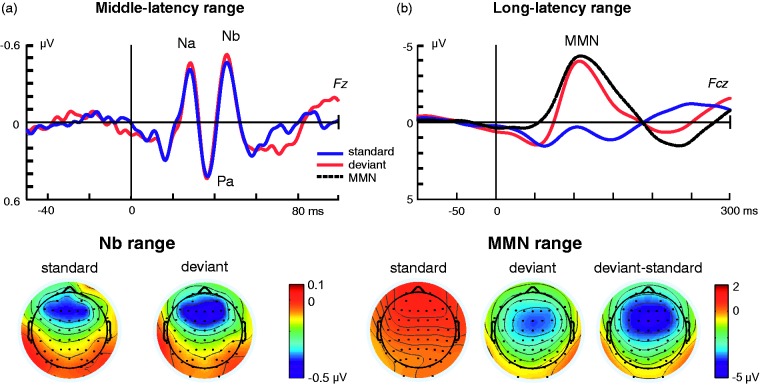
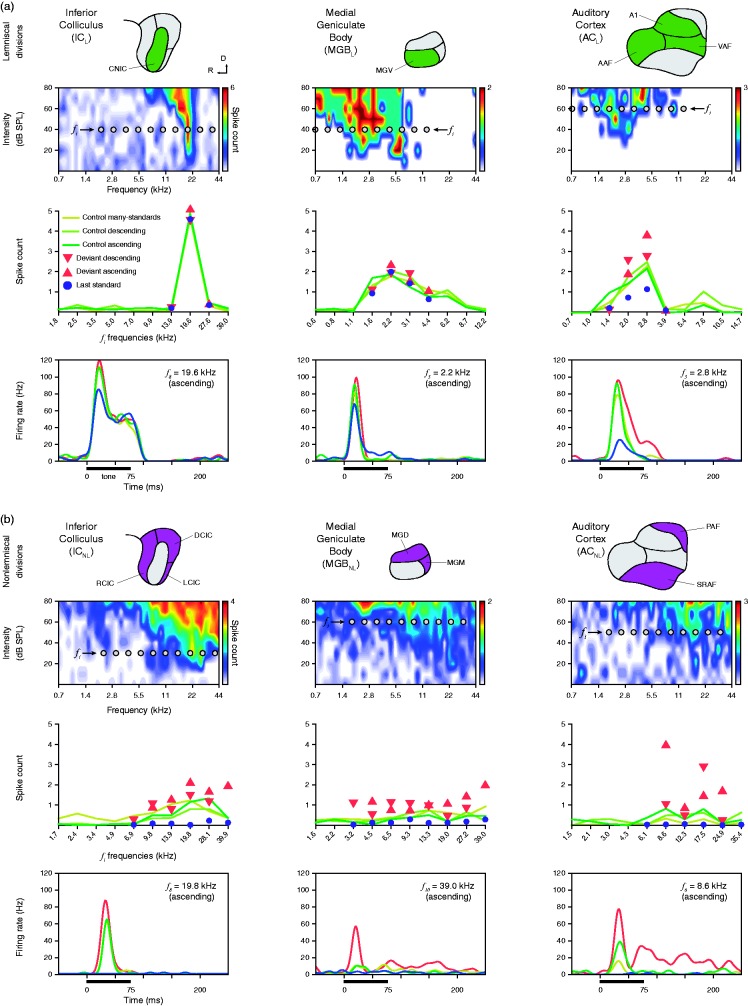
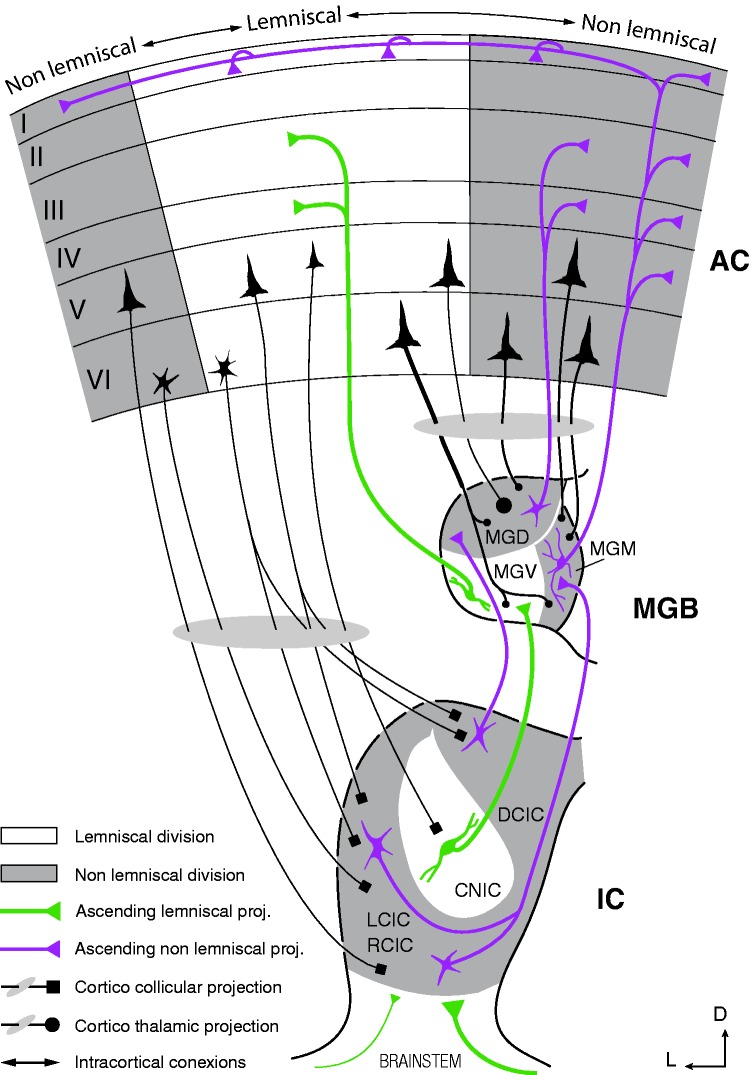
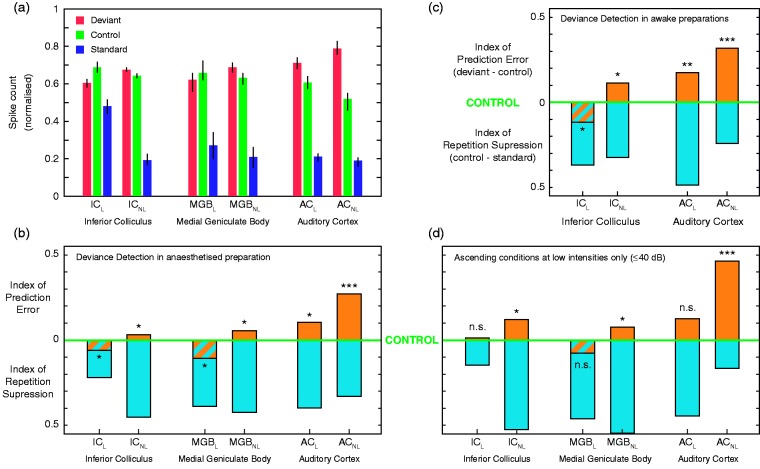
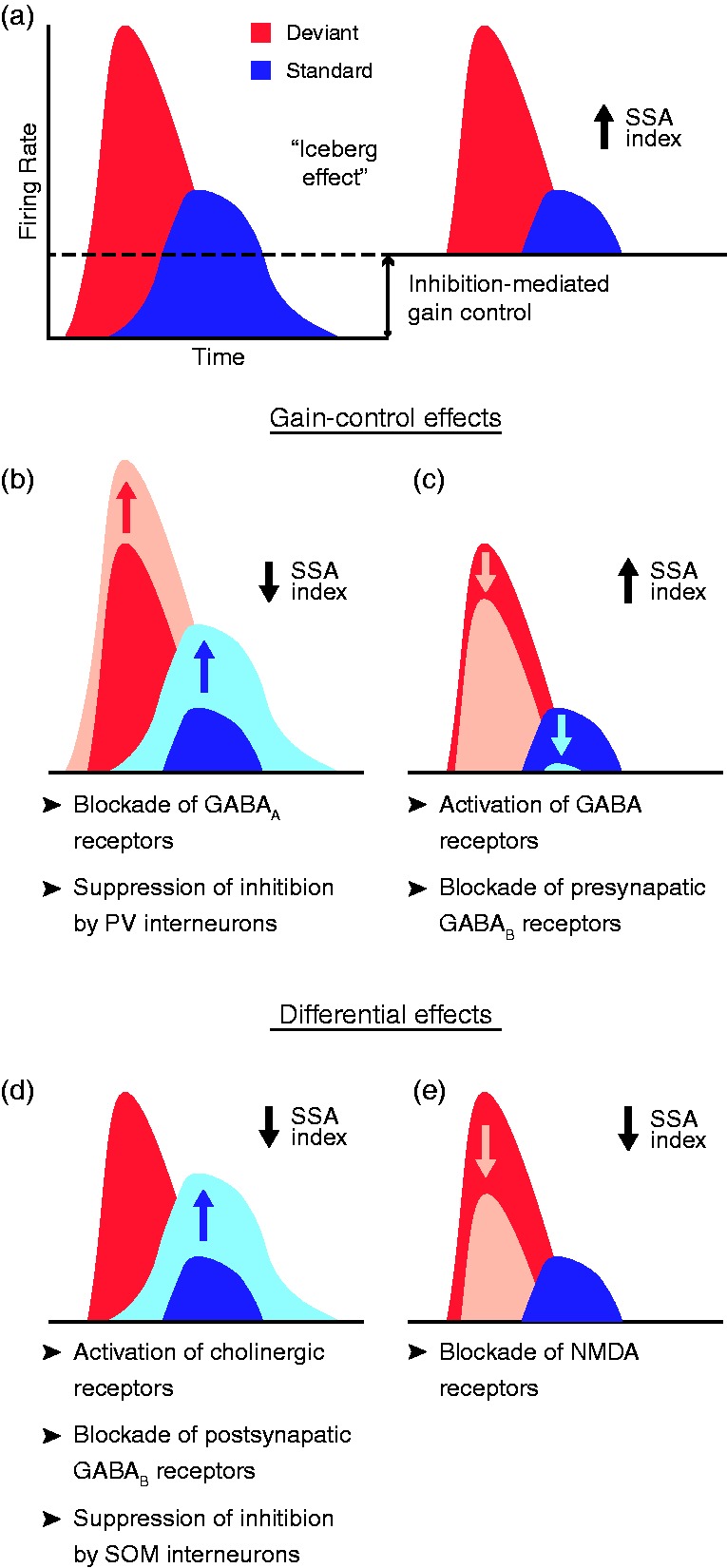
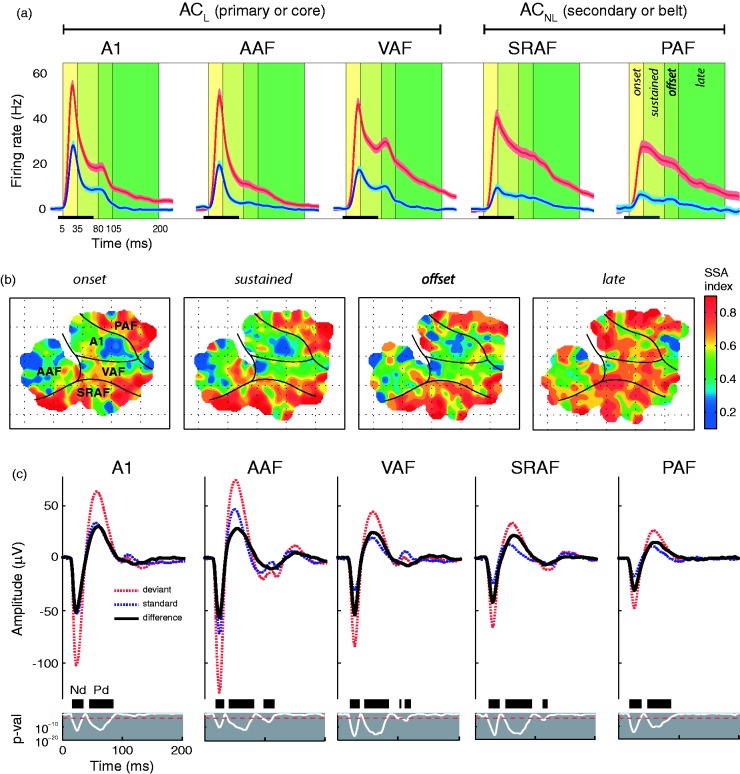
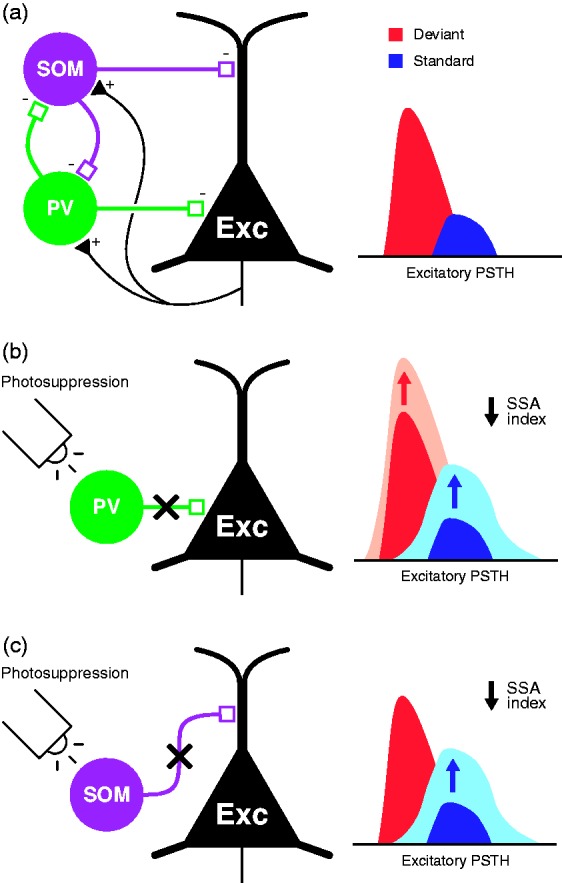
In this review, we attempt to integrate the empirical evidence regarding stimulus-specific adaptation (SSA) and mismatch negativity (MMN) under a predictive coding perspective (also known as Bayesian or hierarchical-inference model). We propose a renewed methodology for SSA study, which enables a further decomposition of deviance detection into repetition suppression and prediction error, thanks to the use of two controls previously introduced in MMN research: the many-standards and the cascade sequences. Focusing on data obtained with cellular recordings, we explain how deviance detection and prediction error are generated throughout hierarchical levels of processing, following two vectors of increasing computational complexity and abstraction along the auditory neuraxis: from subcortical toward cortical stations and from lemniscal toward nonlemniscal divisions. Then, we delve into the particular characteristics and contributions of subcortical and cortical structures to this generative mechanism of hierarchical inference, analyzing what is known about the role of neuromodulation and local microcircuitry in the emergence of mismatch signals. Finally, we describe how SSA and MMN are occurring at similar time frame and cortical locations, and both are affected by the manipulation of N-methyl- D-aspartate receptors. We conclude that there is enough empirical evidence to consider SSA and MMN, respectively, as the microscopic and macroscopic manifestations of the same physiological mechanism of deviance detection in the auditory cortex. Hence, the development of a common theoretical framework for SSA and MMN is all the more recommendable for future studies. In this regard, we suggest a shared nomenclature based on the predictive coding interpretation of deviance detection.
Keywords: MMN; SSA; deviance detection; predictive coding; repetition suppression.
Figures
References
-
- Abolafia J. M., Vergara R., Arnold M. M., Reig R., Sanchez-Vives M. V. (2011) Cortical auditory adaptation in the awake rat and the role of potassium currents. Cerebral Cortex 21(5): 977–990. doi:10.1093/cercor/bhq163. - PubMed
-
- Alain C., Woods D. L., Ogawa K. H. (1994) Brain indices of automatic pattern processing. NeuroReport 6(1): 140–144. doi:10.1097/00001756-199412300-00036. - PubMed
-
- Alho K. (1995) Cerebral generators of mismatch negativity (MMN) and its magnetic counterpart (MMNm) elicited by sound changes. Ear and Hearing 16(1): 38–51. - PubMed
-
- Althen H., Grimm S., Escera C. (2013) Simple and complex acoustic regularities are encoded at different levels of the auditory hierarchy. European Journal of Neuroscience 38(10): 3448–3455. doi:10.1111/ejn.12346. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
