

Transcription Factor Zhx2 Deficiency Reduces Atherosclerosis and Promotes Macrophage Apoptosis in Mice
- PMID: 30026271
- PMCID: PMC6202168
- DOI: 10.1161/ATVBAHA.118.311266
Transcription Factor Zhx2 Deficiency Reduces Atherosclerosis and Promotes Macrophage Apoptosis in Mice
Abstract
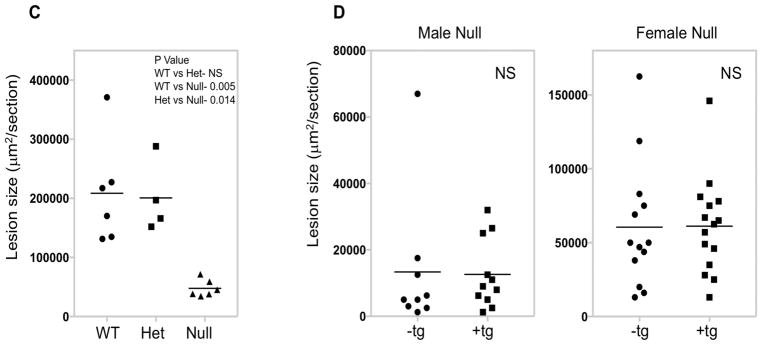
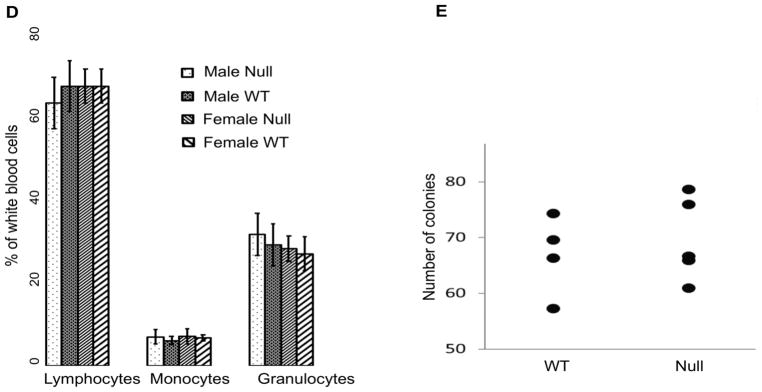
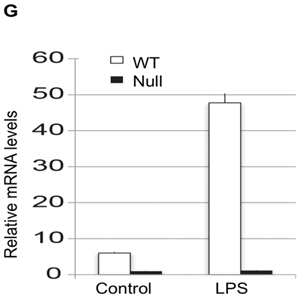
Objective- The objective of this study was to determine the basis of resistance to atherosclerosis of inbred mouse strain BALB/cJ. Approach and Results- BALB/cJ mice carry a naturally occurring null mutation of the gene encoding the transcription factor Zhx2, and genetic analyses suggested that this may confer resistance to atherosclerosis. On a hyperlipidemic low-density lipoprotein receptor null background, BALB/cJ mice carrying the mutant allele for Zhx2 exhibited up to a 10-fold reduction in lesion size as compared with an isogenic strain carrying the wild-type allele. Several lines of evidence, including bone marrow transplantation studies, indicate that this effect of Zhx2 is mediated, in part, by monocytes/macrophages although nonbone marrow-derived pathways are clearly involved as well. Both in culture and in atherosclerotic lesions, macrophages from Zhx2 null mice exhibited substantially increased apoptosis. Zhx2 null macrophages were also enriched for M2 markers. Effects of Zhx2 on proliferation and other bone marrow-derived cells, such as lymphocytes, were at most modest. Expression microarray analyses identified >1000 differentially expressed transcripts between Zhx2 wild-type and null macrophages. To identify the global targets of Zhx2, we performed ChIP-seq (chromatin immunoprecipitation sequencing) studies with the macrophage cell line RAW264.7. The ChIP-seq peaks overlapped significantly with gene expression and together suggested roles for transcriptional repression and apoptosis. Conclusions- A mutation of Zhx2 carried in BALB/cJ mice is responsible in large part for its relative resistance to atherosclerosis. Our results indicate that Zhx2 promotes macrophage survival and proinflammatory functions in atherosclerotic lesions, thereby contributing to lesion growth.
Keywords: apoptosis; atherosclerosis; bone marrow; genetics; macrophages; mice; transcription factor.
Figures








References
-
- Gu L, Johnson MW, Lusis AJ. Quantitative trait locus analysis of plasma lipoprotein levels in an autoimmune mouse model : interactions between lipoprotein metabolism, autoimmune disease, and atherogenesis. Arterioscler Thromb Vasc Biol. 1999;19:442–453. - PubMed
-
- Wang X, Gargalovic P, Wong J, Gu JL, Wu X, Qi H, Wen P, Xi L, Tan B, Gogliotti R, Castellani LW, Chatterjee A, Lusis AJ. Hyplip2, a new gene for combined hyperlipidemia and increased atherosclerosis. Arterioscler Thromb Vasc Biol. 2004;24:1928–1934. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
