

Extending the Time Domain of Neuronal Silencing with Cryptophyte Anion Channelrhodopsins
- PMID: 30027111
- PMCID: PMC6051594
- DOI: 10.1523/ENEURO.0174-18.2018
Extending the Time Domain of Neuronal Silencing with Cryptophyte Anion Channelrhodopsins
Abstract
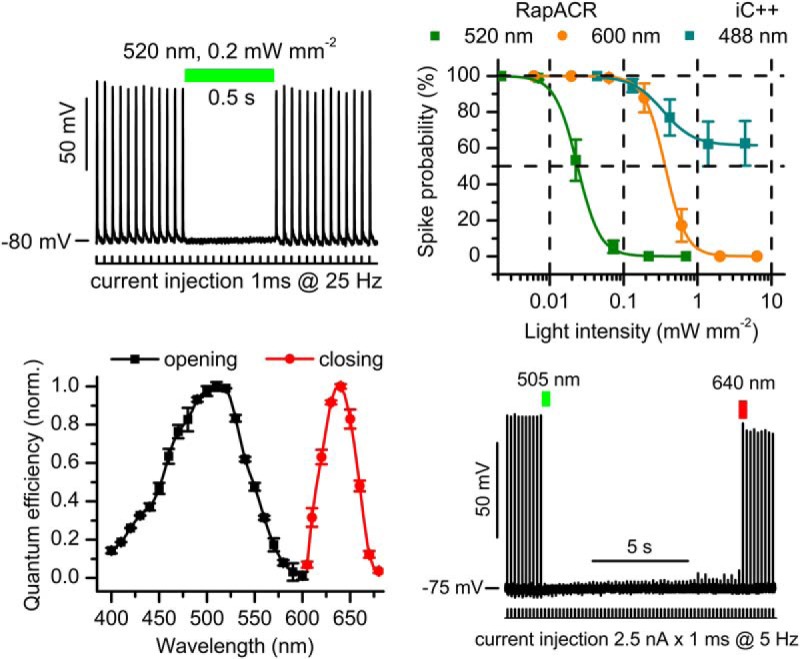
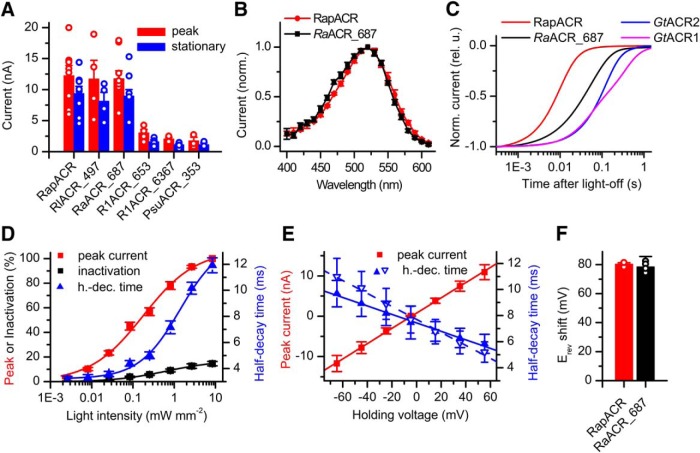
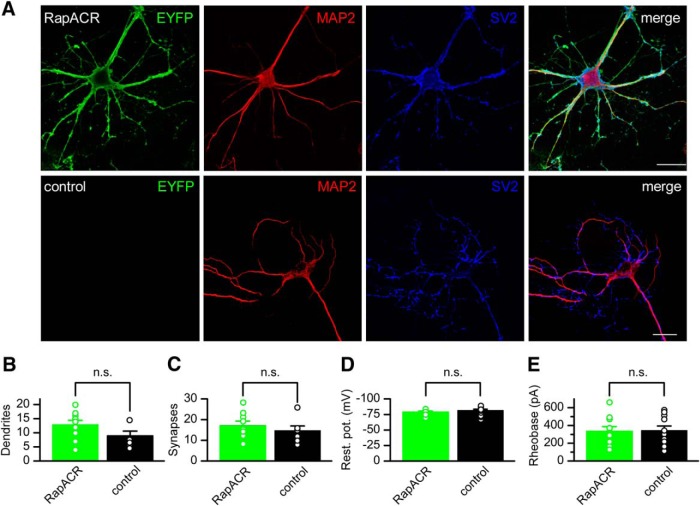
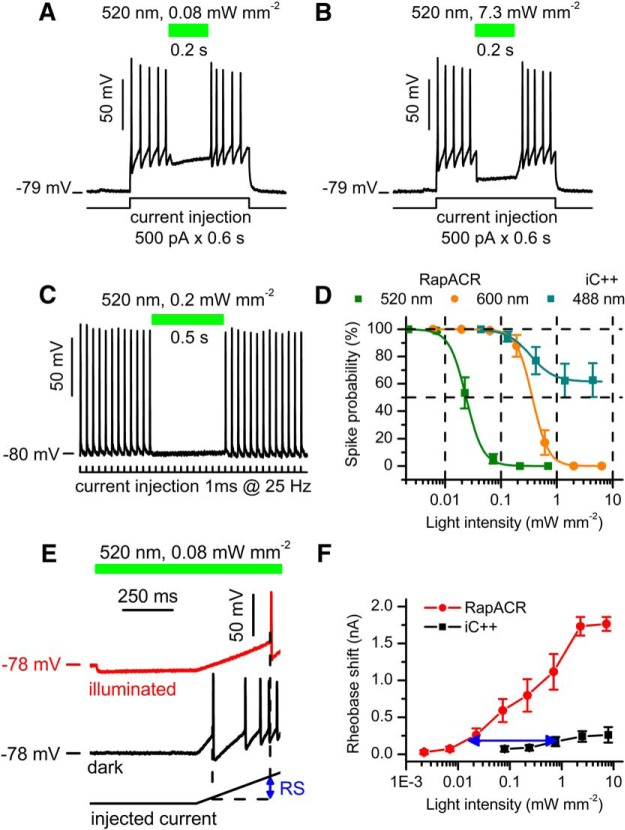
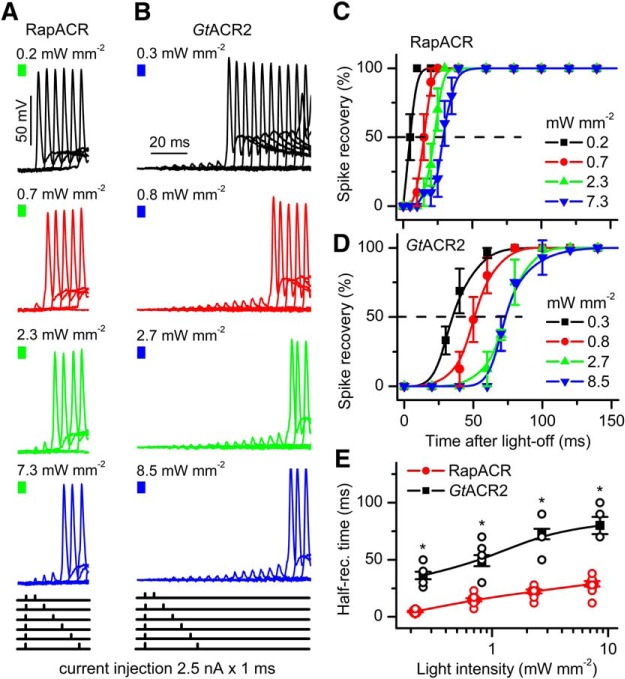
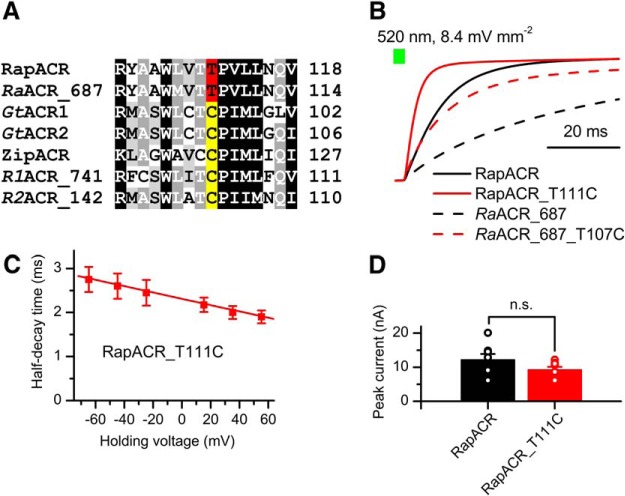
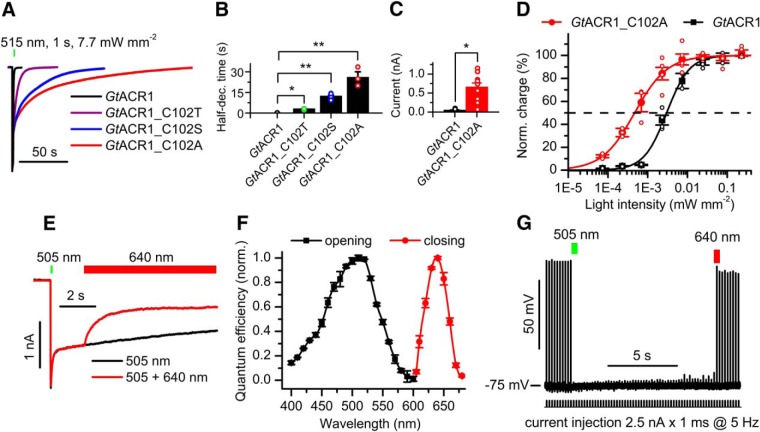
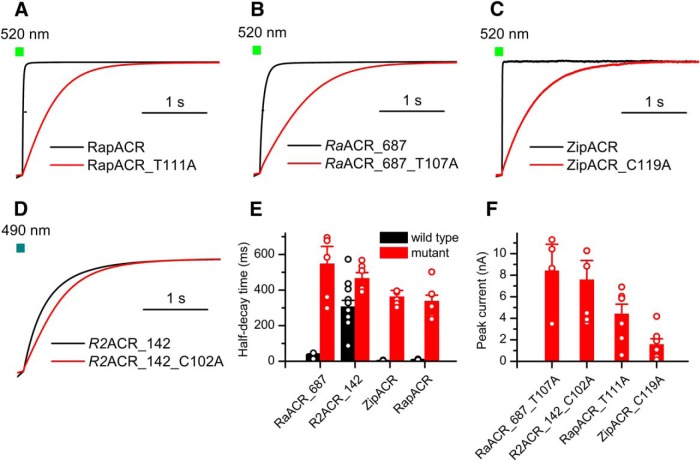
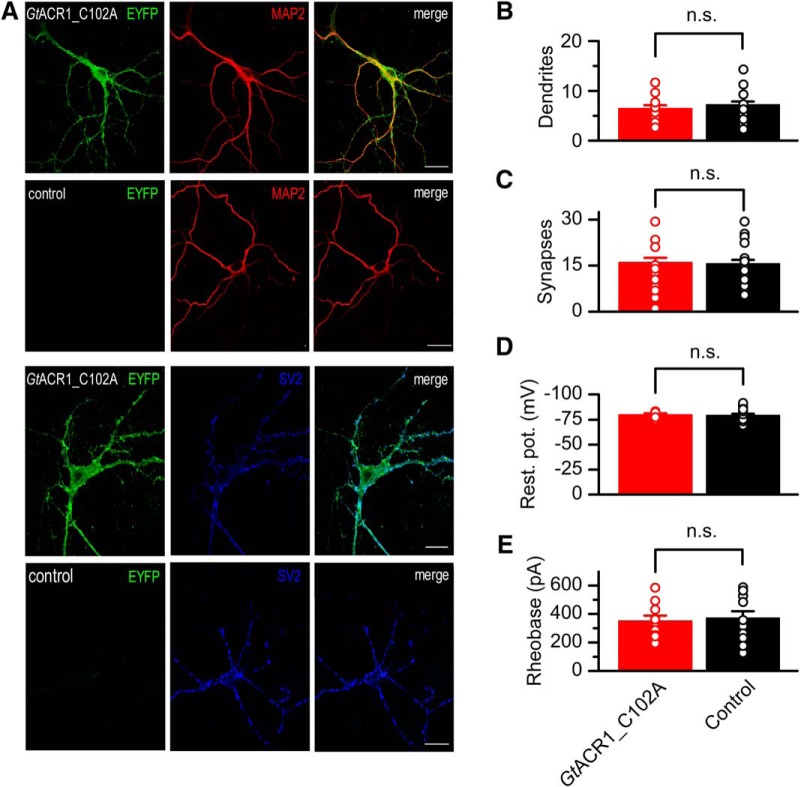
Optogenetic inhibition of specific neuronal types in the brain enables analysis of neural circuitry and is promising for the treatment of a number of neurological disorders. Anion channelrhodopsins (ACRs) from the cryptophyte alga Guillardia theta generate larger photocurrents than other available inhibitory optogenetic tools, but more rapid channels are needed for temporally precise inhibition, such as single-spike suppression, of high-frequency firing neurons. Faster ACRs have been reported, but their potential advantages for time-resolved inhibitory optogenetics have not so far been verified in neurons. We report RapACR, nicknamed so for "rapid," an ACR from Rhodomonas salina, that exhibits channel half-closing times below 10 ms and achieves equivalent inhibition at 50-fold lower light intensity in lentivirally transduced cultured mouse hippocampal neurons as the second-generation engineered Cl--conducting channelrhodopsin iC++. The upper limit of the time resolution of neuronal silencing with RapACR determined by measuring the dependence of spiking recovery after photoinhibition on the light intensity was calculated to be 100 Hz, whereas that with the faster of the two G. theta ACRs was 13 Hz. Further acceleration of RapACR channel kinetics was achieved by site-directed mutagenesis of a single residue in transmembrane helix 3 (Thr111 to Cys). We also show that mutation of another ACR (Cys to Ala at the same position) with a greatly extended lifetime of the channel open state acts as a bistable photochromic tool in mammalian neurons. These molecules extend the time domain of optogenetic neuronal silencing while retaining the high light sensitivity of Guillardia ACRs.
Keywords: channelrhodopsins; chloride ion channels; neuronal inhibition; optogenetics.
Figures
References
-
- Barry PH (1994) JPCalc, a software package for calculating liquid junction potential corrections in patch-clamp, intracellular, epithelial and bilayer measurements and for correcting junction potential measurements. J Neurosci Methods 51:107–116. - PubMed
-
- Bergs A, Schultheis C, Fischer E, Tsunoda SP, Erbguth K, Husson SJ, Govorunova EG, Spudich JL, Nagel G, Gottschalk A, Liewald JF (2018) Rhodopsin optogenetic toolbox v2.0 for light-sensitive excitation and inhibition in Caenorhabditis elegans . PLoS One 13:e0191802. 10.1371/journal.pone.0191802 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials