

Loss of lipin 1-mediated phosphatidic acid phosphohydrolase activity in muscle leads to skeletal myopathy in mice
- PMID: 30028636
- PMCID: PMC6355067
- DOI: 10.1096/fj.201800361R
Loss of lipin 1-mediated phosphatidic acid phosphohydrolase activity in muscle leads to skeletal myopathy in mice
Abstract
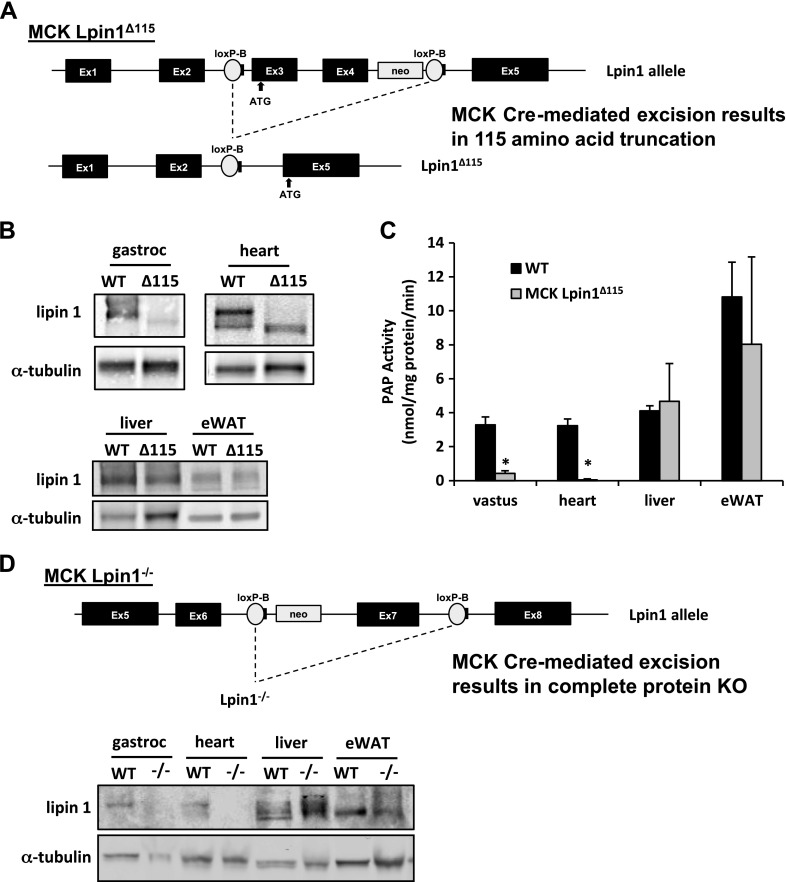
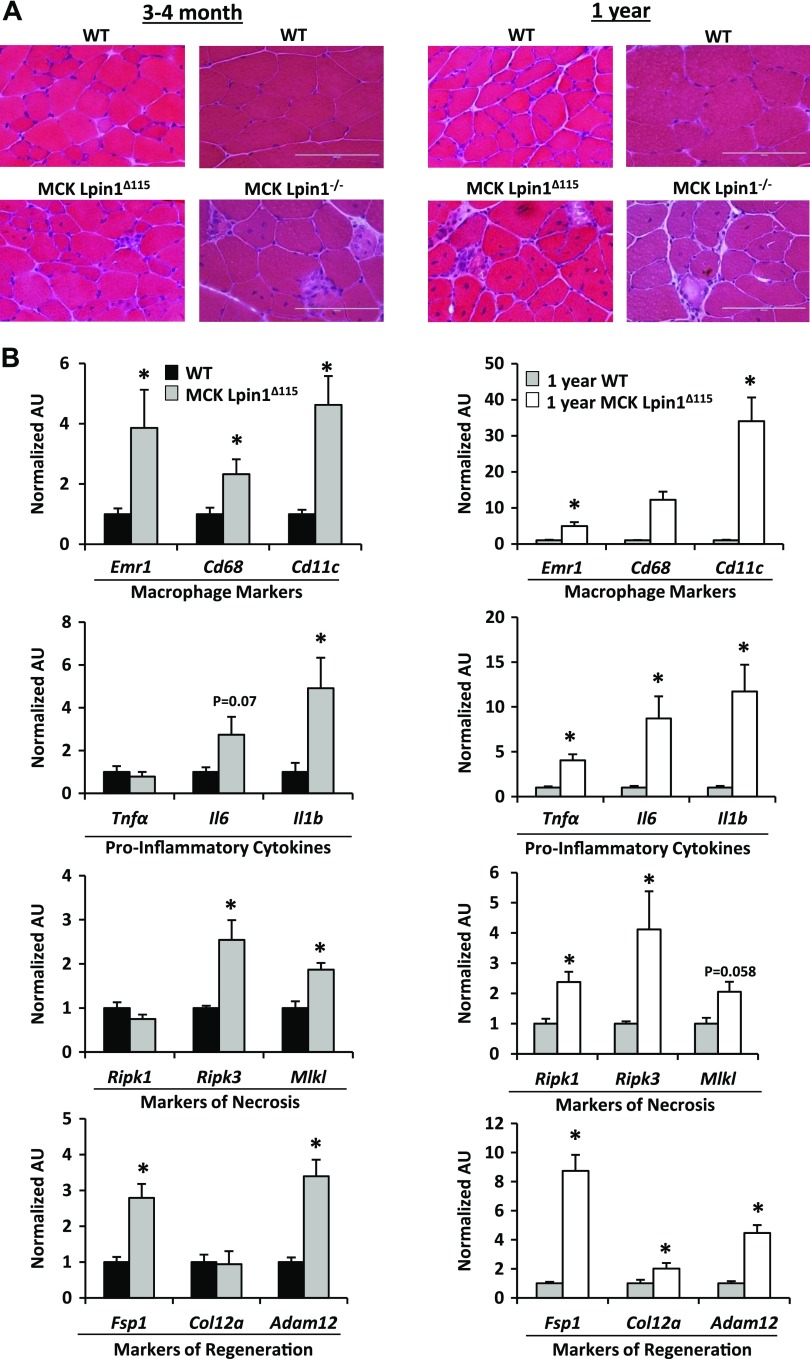
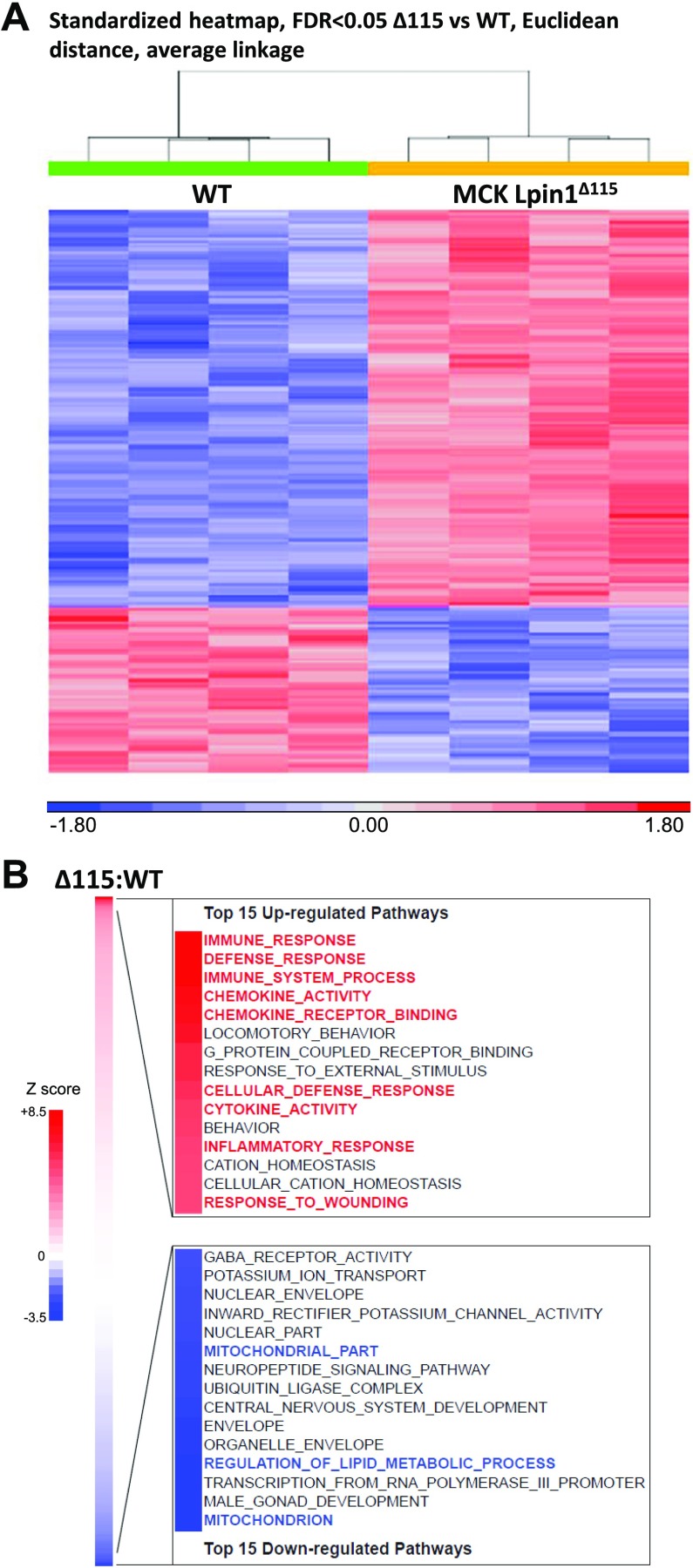
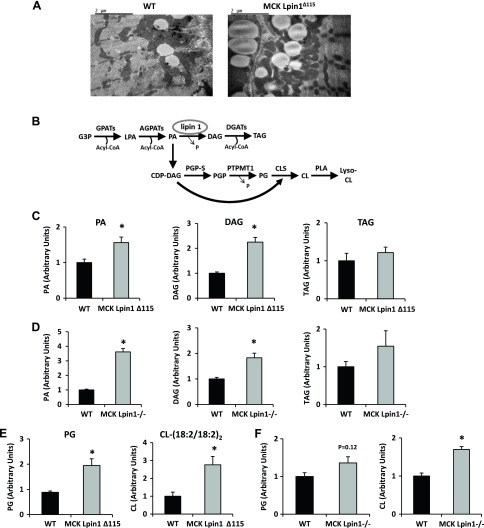
Lipin 1 regulates glycerolipid homeostasis by acting as a phosphatidic acid phosphohydrolase (PAP) enzyme in the triglyceride-synthesis pathway and by regulating transcription factor activity. Mutations in human lipin 1 are a common cause of recurrent rhabdomyolysis in children. Mice with constitutive whole-body lipin 1 deficiency have been used to examine mechanisms connecting lipin 1 deficiency to myocyte injury. However, that mouse model is confounded by lipodystrophy not phenocopied in people. Herein, 2 muscle-specific mouse models were studied: 1) Lpin1 exon 3 and 4 deletion, resulting in a hypomorphic protein without PAP activity, but which preserved transcriptional coregulatory function; and 2) Lpin1 exon 7 deletion, resulting in total protein loss. In both models, skeletal muscles exhibited a chronic myopathy with ongoing muscle fiber necrosis and regeneration and accumulation of phosphatidic acid and, paradoxically, diacylglycerol. Additionally, lipin 1-deficient mice had abundant, but abnormal, mitochondria likely because of impaired autophagy. Finally, these mice exhibited increased plasma creatine kinase following exhaustive exercise when unfed. These data suggest that mice lacking lipin 1-mediated PAP activity in skeletal muscle may serve as a model for determining the mechanisms by which lipin 1 deficiency leads to myocyte injury and for testing potential therapeutic approaches.-Schweitzer, G. G., Collier, S. L., Chen, Z., McCommis, K. S., Pittman, S. K., Yoshino, J., Matkovich, S. J., Hsu, F.-F., Chrast, R., Eaton, J. M., Harris, T. E., Weihl, C. C., Finck, B. N. Loss of lipin 1-mediated phosphatidic acid phosphohydrolase activity in muscle leads to skeletal myopathy in mice.
Keywords: LPIN1; autophagy; diacylglycerol; rhabdomyolysis; triacylglycerol.
Conflict of interest statement
The authors thank Dr. Alan Pestronk and Dr. Abhinav Diwan for technical assistance in interpreting histology, and Dennis J. Dietzen for determining plasma creatine kinase (all from Washington University). This work was supported by U.S. National Institutes of Health (NIH)
Figures
References
-
- Peterson T. R., Sengupta S. S., Harris T. E., Carmack A. E., Kang S. A., Balderas E., Guertin D. A., Madden K. L., Carpenter A. E., Finck B. N., Sabatini D. M. (2011) mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell 146, 408–420 10.1016/j.cell.2011.06.034 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK101946/DK/NIDDK NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- R01 HL119225/HL/NHLBI NIH HHS/United States
- P30 CA091842/CA/NCI NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- R01 AG031867/AG/NIA NIH HHS/United States
- KL2 TR000450/TR/NCATS NIH HHS/United States
- T32 HL007275/HL/NHLBI NIH HHS/United States
- T32 HL007081/HL/NHLBI NIH HHS/United States
- R01 DK078187/DK/NIDDK NIH HHS/United States
- P41 GM103422/GM/NIGMS NIH HHS/United States
- R01 DK104995/DK/NIDDK NIH HHS/United States
- KL2 TR002346/TR/NCATS NIH HHS/United States
- UL1 TR000448/TR/NCATS NIH HHS/United States
- T32 DK007120/DK/NIDDK NIH HHS/United States
- P60 DK020579/DK/NIDDK NIH HHS/United States
- T32 GM007055/GM/NIGMS NIH HHS/United States
- UL1 TR002345/TR/NCATS NIH HHS/United States
- K24 AR073317/AR/NIAMS NIH HHS/United States
- R01 AR068797/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
