

Mechanistic Insights into Colorectal Cancer Phenomics from Fundamental and Organotypic Model Studies
- PMID: 30028958
- PMCID: PMC6240511
- DOI: 10.1016/j.ajpath.2018.05.021
Mechanistic Insights into Colorectal Cancer Phenomics from Fundamental and Organotypic Model Studies
Abstract
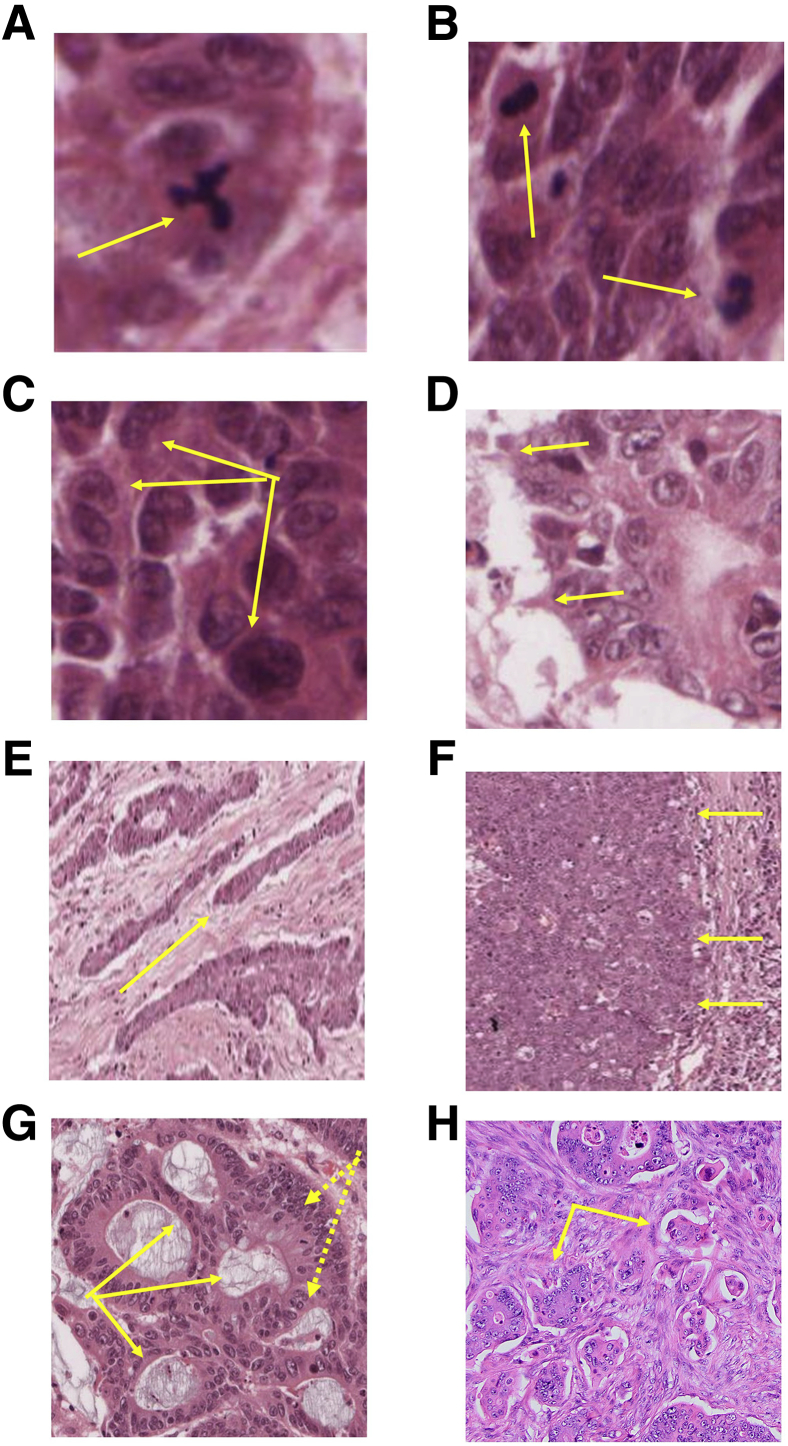
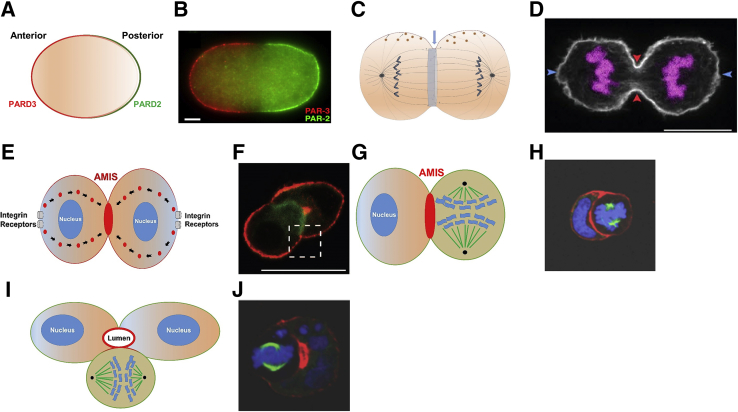
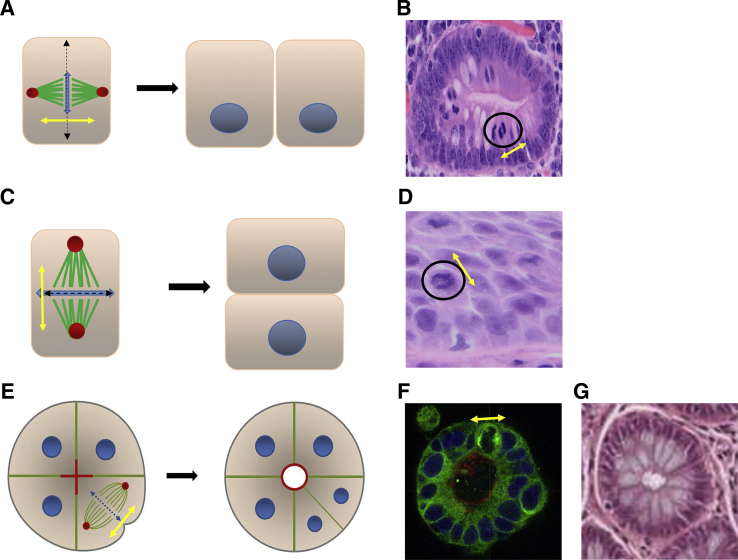
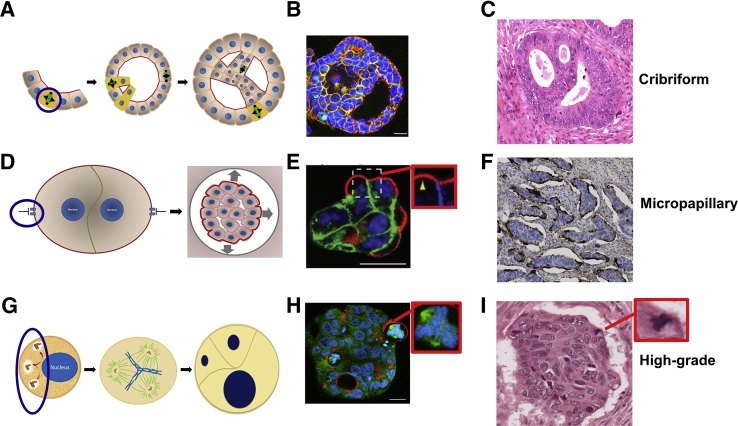
Colorectal cancer (CRC) diagnosis and prognostic stratification are based on histopathologic assessment of cell or nuclear pleomorphism, aberrant mitotic figures, altered glandular architecture, and other phenomic abnormalities. This complexity is driven by oncogenic perturbation of tightly coordinated spatiotemporal signaling to disrupt multiple scales of tissue organization. This review clarifies molecular and cellular mechanisms underlying common CRC histologic features and helps understand how the CRC genome controls core aspects of tumor aggressiveness. It further explores a spatiotemporal framework for CRC phenomics based on regulation of living cells in fundamental and organotypic model systems. The review also discusses tissue homeostasis, considers distinct classes of oncogenic perturbations, and evolution of cellular or multicellular cancer phenotypes. It further explores the molecular controls of cribriform, micropapillary, and high-grade CRC morphology in organotypic culture models and assesses relevant translational studies. In addition, the review delves into complexities of morphologic plasticity whereby a single molecular signature generates heterogeneous cancer phenotypes, and, conversely, morphologically homogeneous tumors show substantive molecular diversity. Principles outlined may aid mechanistic interpretation of omics data in a setting of cancer pathology, provide insight into CRC consensus molecular subtypes, and better define principles for CRC prognostic stratification.
Copyright © 2018 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Heng Y.J., Lester S.C., Tse G.M., Factor R.E., Allison K.H., Collins L.C., Chen Y.Y., Jensen K.C., Johnson N.B., Jeong J.C., Punjabi R., Shin S.J., Singh K., Krings G., Eberhard D.A., Tan P.H., Korski K., Waldman F.M., Gutman D.A., Sanders M., Reis-Filho J.S., Flanagan S.R., Gendoo D.M., Chen G.M., Haibe-Kains B., Ciriello G., Hoadley K.A., Perou C.M., Beck A.H. The molecular basis of breast cancer pathological phenotypes. J Pathol. 2017;241:375–391. - PMC - PubMed
-
- Hoadley K.A., Yau C., Hinoue T., Wolf D.M., Lazar A.J., Drill E., Shen R., Taylor A.M., Cherniack A.D., Thorsson V., Akbani R., Bowlby R., Wong C.K., Wiznerowicz M., Sanchez-Vega F., Robertson A.G., Schneider B.G., Lawrence M.S., Noushmehr H., Malta T.M., Cancer Genome Atlas Network. Stuart J.M., Benz C.C., Laird P.W. Cell-of-origin patterns dominate the molecular classification of 10,000 tumors from 33 types of cancer. Cell. 2018;173:291–304.e6. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
