

Detecting and interpreting DNA methylation marks
- PMID: 30031306
- PMCID: PMC6322410
- DOI: 10.1016/j.sbi.2018.06.004
Detecting and interpreting DNA methylation marks
Abstract
The generation, alteration, recognition, and erasure of epigenetic modifications of DNA are fundamental to controlling gene expression in mammals. These covalent DNA modifications include cytosine methylation by AdoMet-dependent methyltransferases and 5-methylcytosine oxidation by Fe(II)-dependent and α-ketoglutarate-dependent dioxygenases. Sequence-specific transcription factors are responsible for interpreting the modification status of specific regions of chromatin. This review focuses on recent developments in characterizing the functional and structural links between the modification status of two DNA bases: 5-methylcytosine and 5-methyluracil (thymine).
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest statement
The authors declare no conflict of interest.
Figures
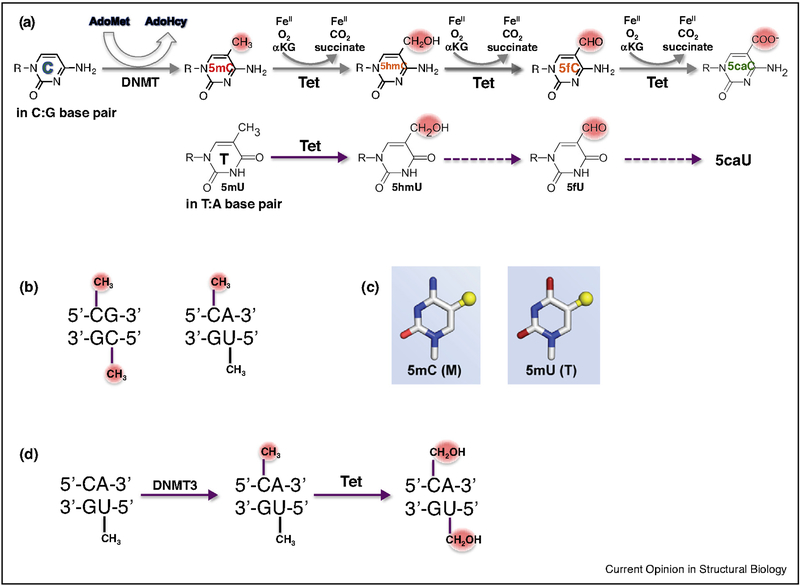
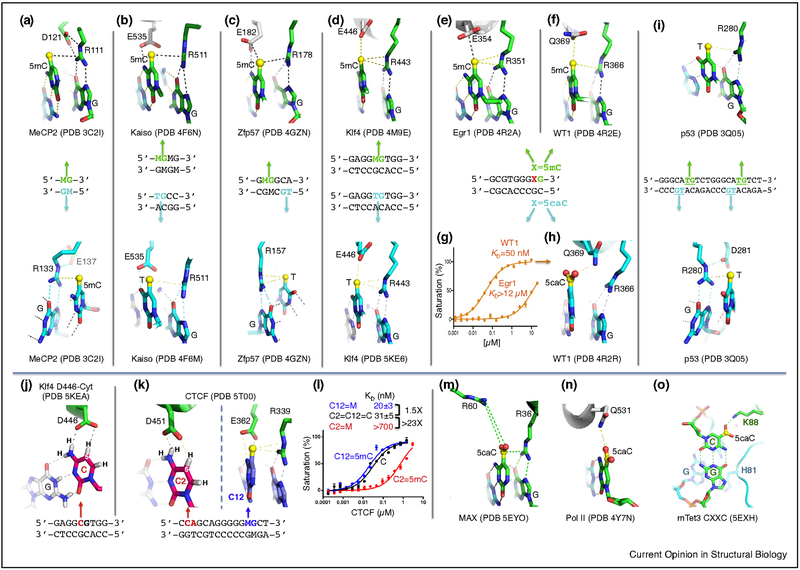
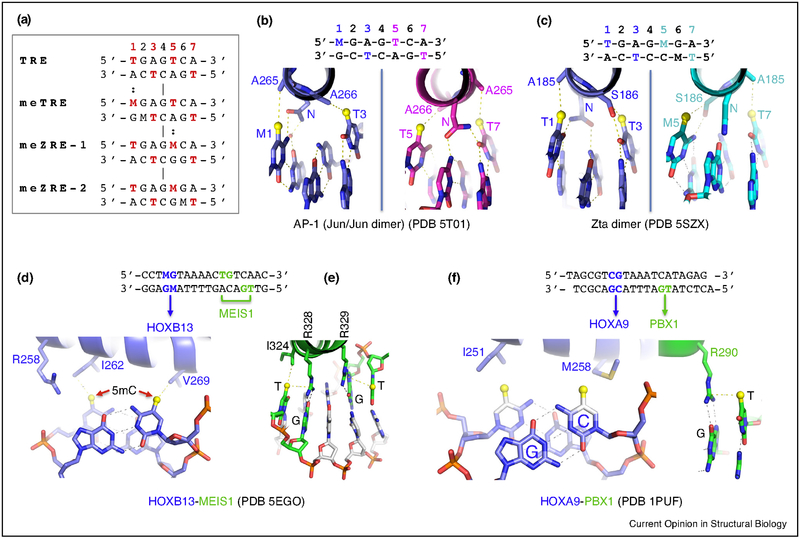
References
-
- Pfaffeneder T, Hackner B, Truss M, Munzel M, Muller M, Deiml CA, Hagemeier C, Carell T: The discovery of 5-formylcytosine in embryonic stem cell DNA. Angew Chem Int Ed Engl 2011, 50:7008–7012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
