

Lifecycle modelling systems support inosine monophosphate dehydrogenase (IMPDH) as a pro-viral factor and antiviral target for New World arenaviruses
- PMID: 30031760
- PMCID: PMC11099991
- DOI: 10.1016/j.antiviral.2018.07.009
Lifecycle modelling systems support inosine monophosphate dehydrogenase (IMPDH) as a pro-viral factor and antiviral target for New World arenaviruses
Abstract
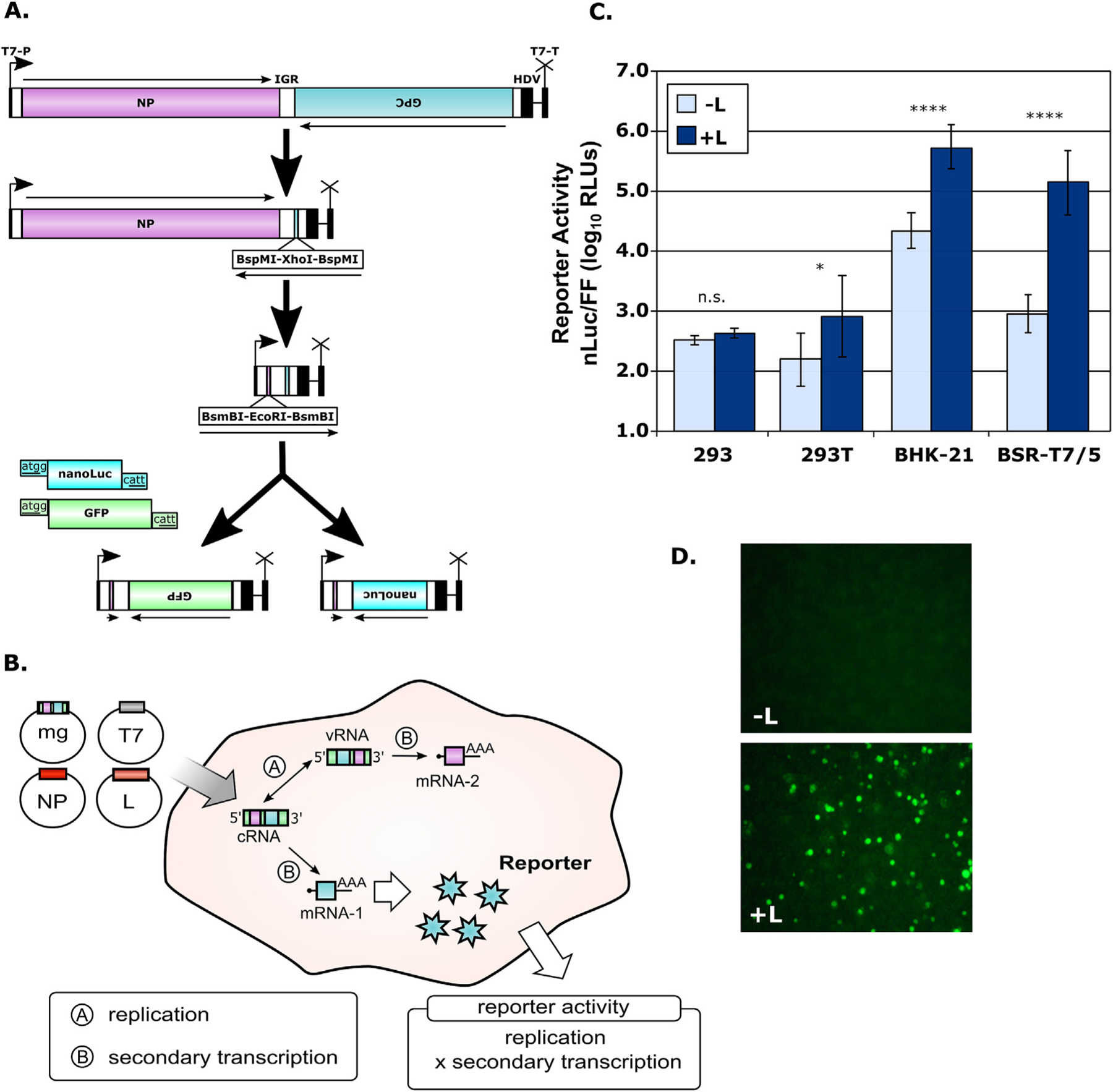
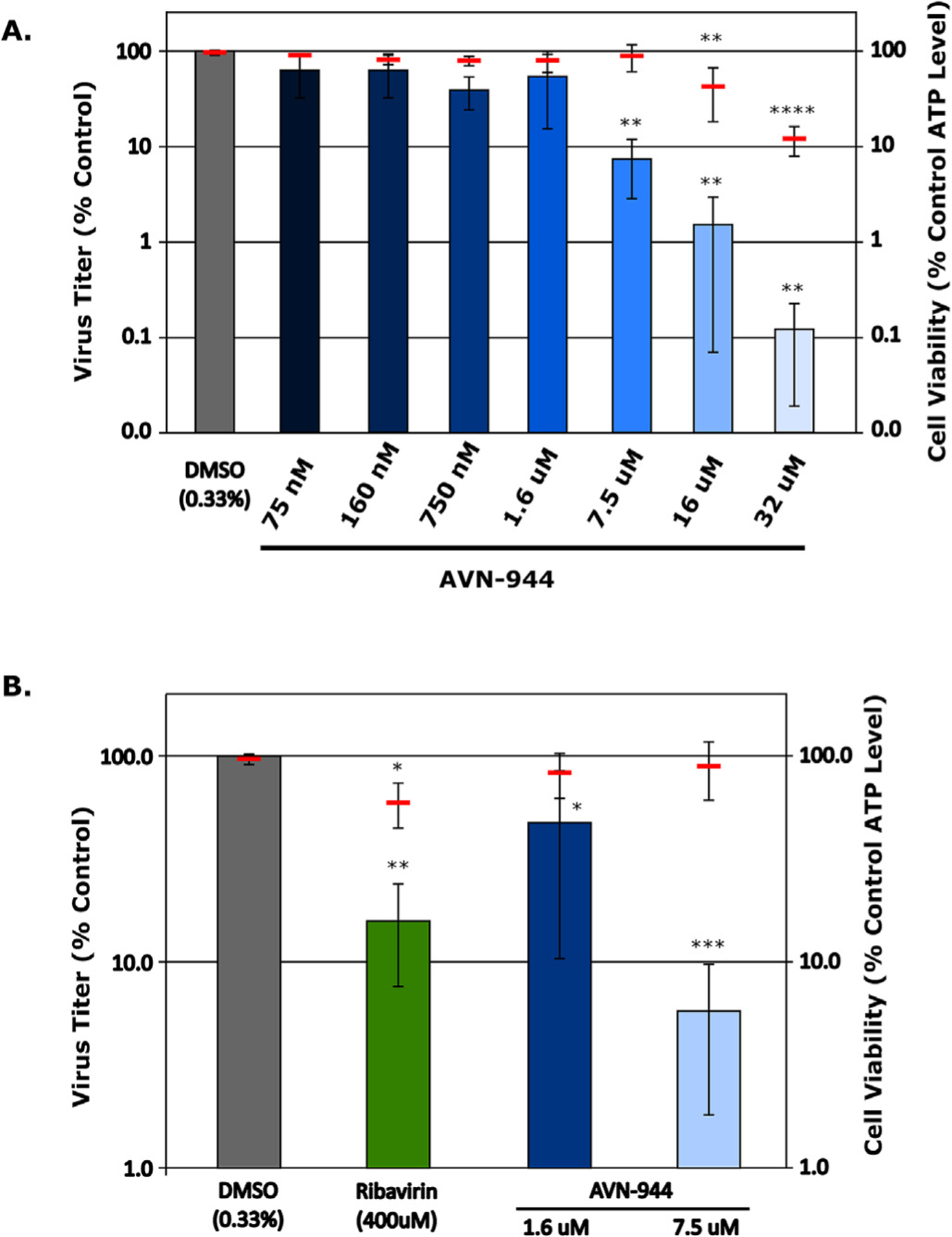
Infection with Junín virus (JUNV) is currently being effectively managed in the endemic region using a combination of targeted vaccination and plasma therapy. However, the long-term sustainability of plasma therapy is unclear and similar resources are not available for other New World arenaviruses. As a result, there has been renewed interest regarding the potential of drug-based therapies. To facilitate work on this issue, we present the establishment and subsequent optimization of a JUNV minigenome system to a degree suitable for high-throughput miniaturization, thereby providing a screening platform focused solely on factors affecting RNA synthesis. Using this tool, we conducted a limited drug library screen and identified AVN-944, a non-competitive inosine monophosphate dehydrogenase (IMPDH) inhibitor, as an inhibitor of arenavirus RNA synthesis. We further developed a transcription and replication competent virus-like particle (trVLP) system based on these minigenomes and used it to screen siRNAs against IMPDH, verifying its role in supporting arenavirus RNA synthesis. The antiviral effect of AVN-944, as well as siRNA inhibition, on JUNV RNA synthesis supports that, despite playing only a minor role in the activity of ribavirin, exclusive IMPDH inhibitors may indeed have significant therapeutic potential for use against New World arenaviruses. Finally, we confirmed that AVN-944 is also active against arenavirus infection in cell culture, supporting the suitability of arenavirus lifecycle modelling systems as tools for the screening and identification, as well as the mechanistic characterization, of novel antiviral compounds.
Keywords: AVN-944; Arenavirus; Inosine monophosphate dehydrogenase (IMPDH); Life cycle modelling systems.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures



References
-
- Borroto-Esoda K, Myrick F, Feng J, Jeffrey J, Furman P, 2004. In vitro combination of amdoxovir and the inosine monophosphate dehydrogenase inhibitors mycophenolic acid and ribavirin demonstrates potent activity against wild-type and drug-resistant variants of human immunodeficiency virus type 1. Antimicrob. Agents Chemother 48, 4387–4394. - PMC - PubMed
-
- Emonet SF, Seregin AV, Yun NE, Poussard AL, Walker AG, de la Torre JC, Paessler S, 2011. Rescue from cloned cDNAs and in vivo characterization of recombinant pathogenic Romero and live-attenuated Candid #1 strains of Junin virus, the causative agent of Argentine hemorrhagic fever disease. J. Virol 85, 1473–1483. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
