

Mosaic-variegated aneuploidy syndrome mutation or haploinsufficiency in Cep57 impairs tumor suppression
- PMID: 30035751
- PMCID: PMC6063474
- DOI: 10.1172/JCI120316
Mosaic-variegated aneuploidy syndrome mutation or haploinsufficiency in Cep57 impairs tumor suppression
Abstract
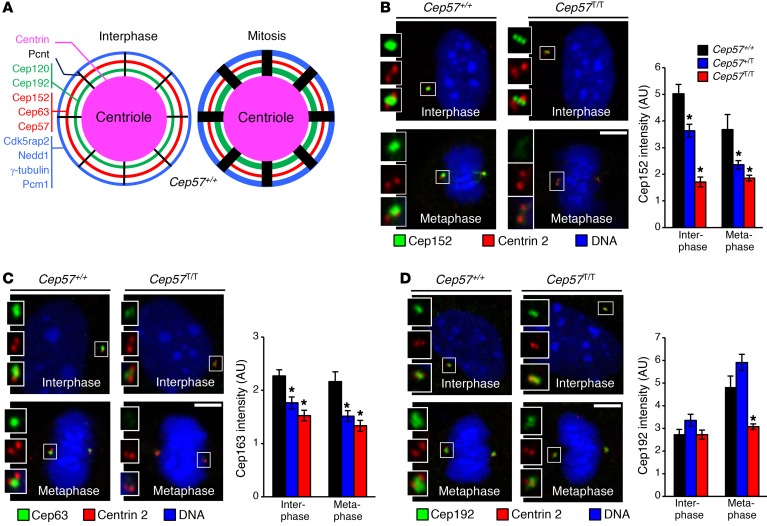
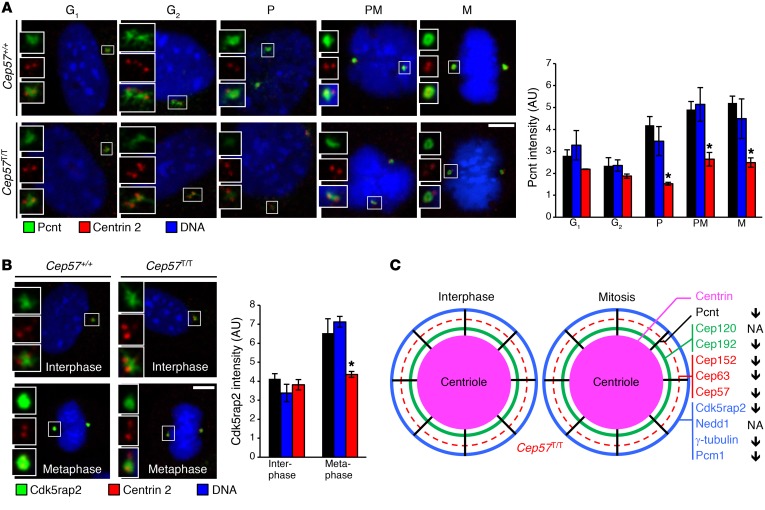
A homozygous truncating frameshift mutation in CEP57 (CEP57T/T) has been identified in a subset of mosaic-variegated aneuploidy (MVA) patients; however, the physiological roles of the centrosome-associated protein CEP57 that contribute to disease are unknown. To investigate these, we have generated a mouse model mimicking this disease mutation. Cep57T/T mice died within 24 hours after birth with short, curly tails and severely impaired vertebral ossification. Osteoblasts in lumbosacral vertebrae of Cep57T/T mice were deficient for Fgf2, a Cep57 binding partner implicated in diverse biological processes, including bone formation. Furthermore, a broad spectrum of tissues of Cep57T/T mice had severe aneuploidy at birth, consistent with the MVA patient phenotype. Cep57T/T mouse embryonic fibroblasts and patient-derived skin fibroblasts failed to undergo centrosome maturation in G2 phase, causing premature centriole disjunction, centrosome amplification, aberrant spindle formation, and high rates of chromosome missegregation. Mice heterozygous for the truncating frameshift mutation or a Cep57-null allele were overtly indistinguishable from WT mice despite reduced Cep57 protein levels, yet prone to aneuploidization and cancer, with tumors lacking evidence for loss of heterozygosity. This study identifies Cep57 as a haploinsufficient tumor suppressor with biologically diverse roles in centrosome maturation and Fgf2-mediated bone formation.
Keywords: Bone development; Cell Biology; Genetic diseases; Oncology.
Conflict of interest statement
Figures







References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
