

Top-down, contextual entrainment of neuronal oscillations in the auditory thalamocortical circuit
- PMID: 30037997
- PMCID: PMC6094129
- DOI: 10.1073/pnas.1714684115
Top-down, contextual entrainment of neuronal oscillations in the auditory thalamocortical circuit
Abstract
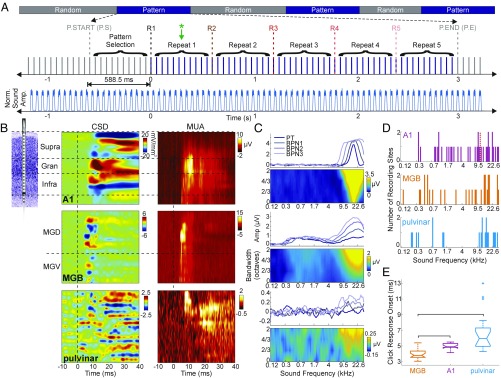
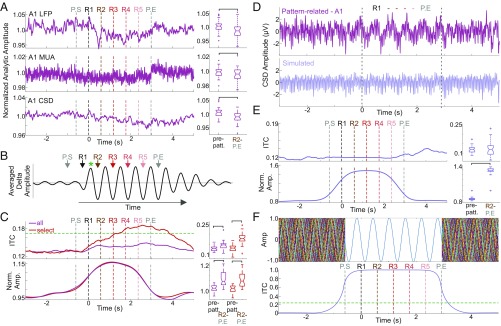
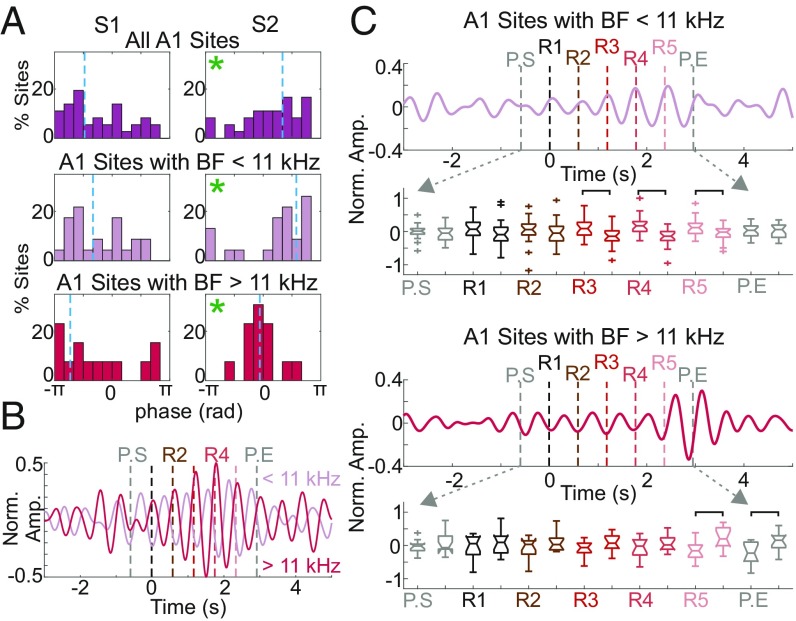
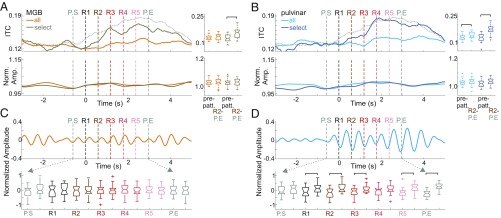
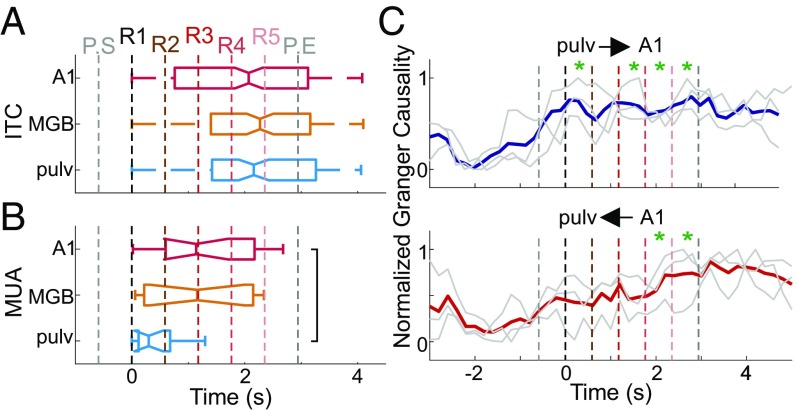
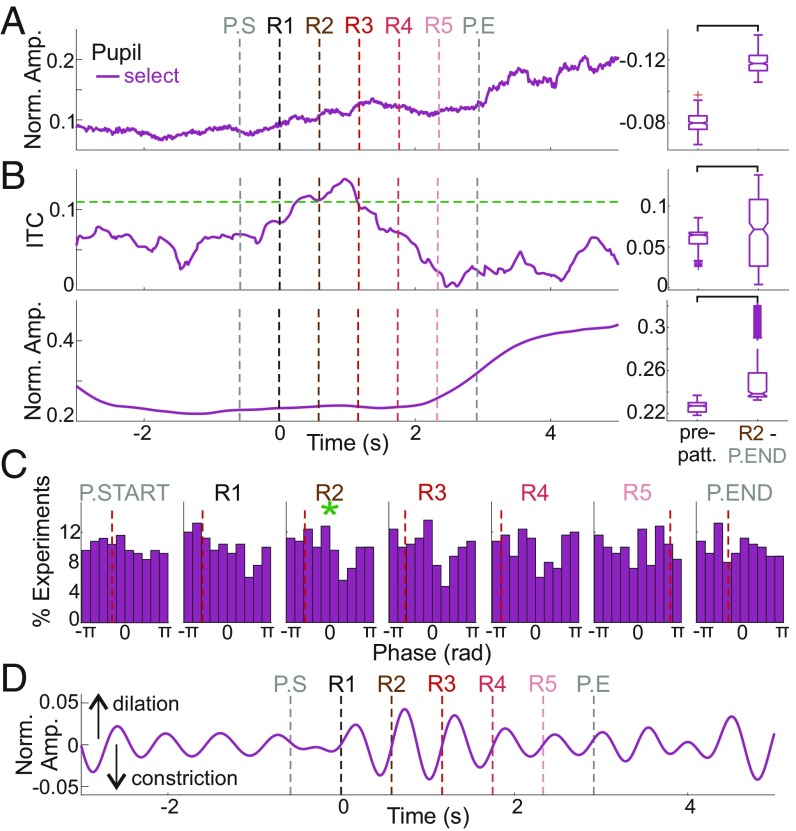
Prior studies have shown that repetitive presentation of acoustic stimuli results in an alignment of ongoing neuronal oscillations to the sequence rhythm via oscillatory entrainment by external cues. Our study aimed to explore the neural correlates of the perceptual parsing and grouping of complex repeating auditory patterns that occur based solely on statistical regularities, or context. Human psychophysical studies suggest that the recognition of novel auditory patterns amid a continuous auditory stimulus sequence occurs automatically halfway through the first repetition. We hypothesized that once repeating patterns were detected by the brain, internal rhythms would become entrained, demarcating the temporal structure of these repetitions despite lacking external cues defining pattern on- or offsets. To examine the neural correlates of pattern perception, neuroelectric activity of primary auditory cortex (A1) and thalamic nuclei was recorded while nonhuman primates passively listened to streams of rapidly presented pure tones and bandpass noise bursts. At arbitrary intervals, random acoustic patterns composed of 11 stimuli were repeated five times without any perturbance of the constant stimulus flow. We found significant delta entrainment by these patterns in the A1, medial geniculate body, and medial pulvinar. In A1 and pulvinar, we observed a statistically significant, pattern structure-aligned modulation of neuronal firing that occurred earliest in the pulvinar, supporting the idea that grouping and detecting complex auditory patterns is a top-down, context-driven process. Besides electrophysiological measures, a pattern-related modulation of pupil diameter verified that, like humans, nonhuman primates consciously detect complex repetitive patterns that lack physical boundaries.
Keywords: auditory patterns; auditory perception; macaque; oscillations; rhythms.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
