

Enteroendocrine cells switch hormone expression along the crypt-to-villus BMP signalling gradient
- PMID: 30038251
- PMCID: PMC6276989
- DOI: 10.1038/s41556-018-0143-y
Enteroendocrine cells switch hormone expression along the crypt-to-villus BMP signalling gradient
Abstract
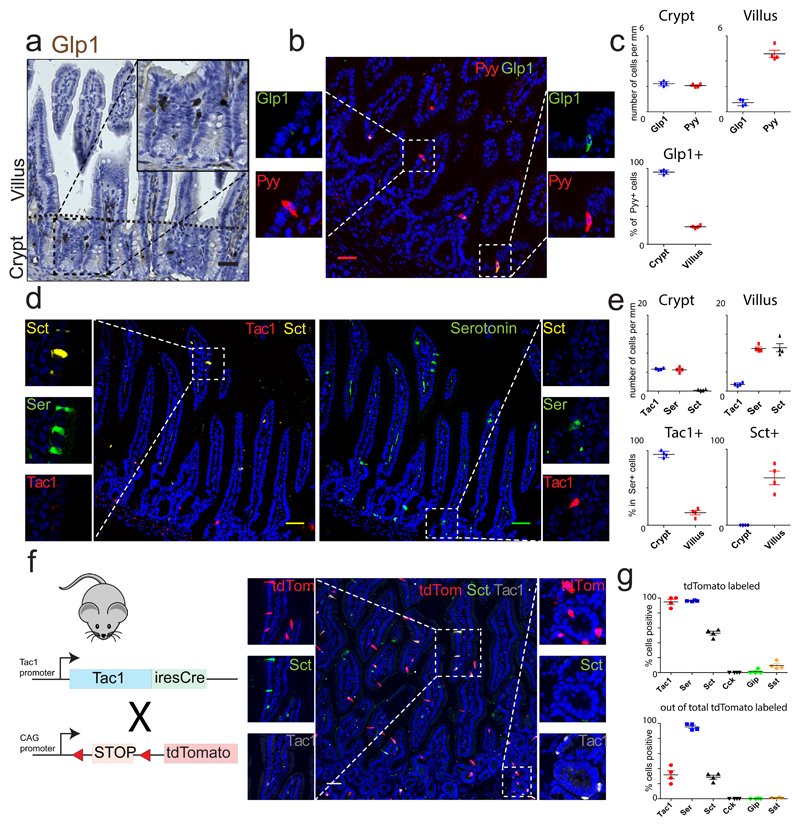
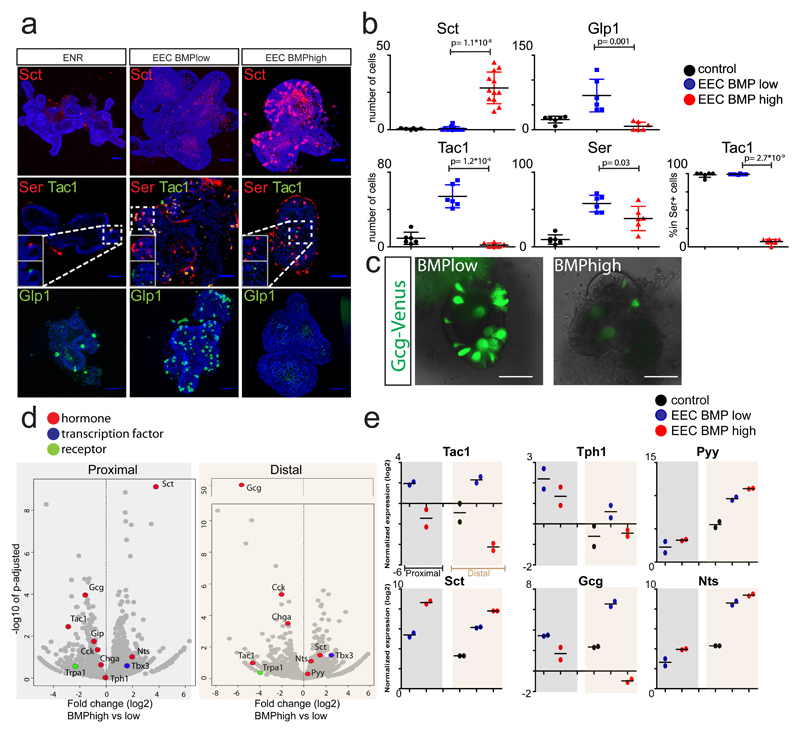
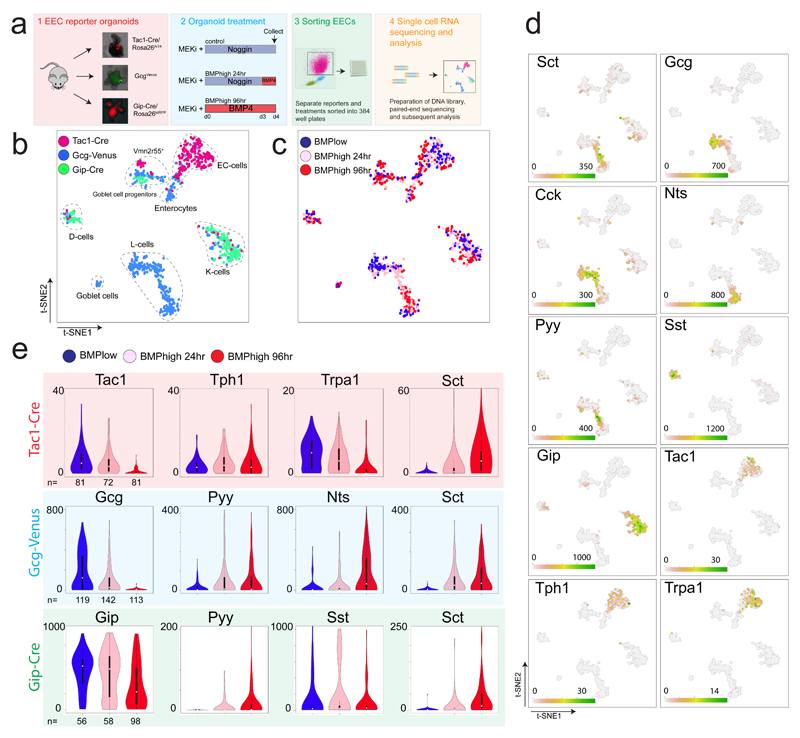
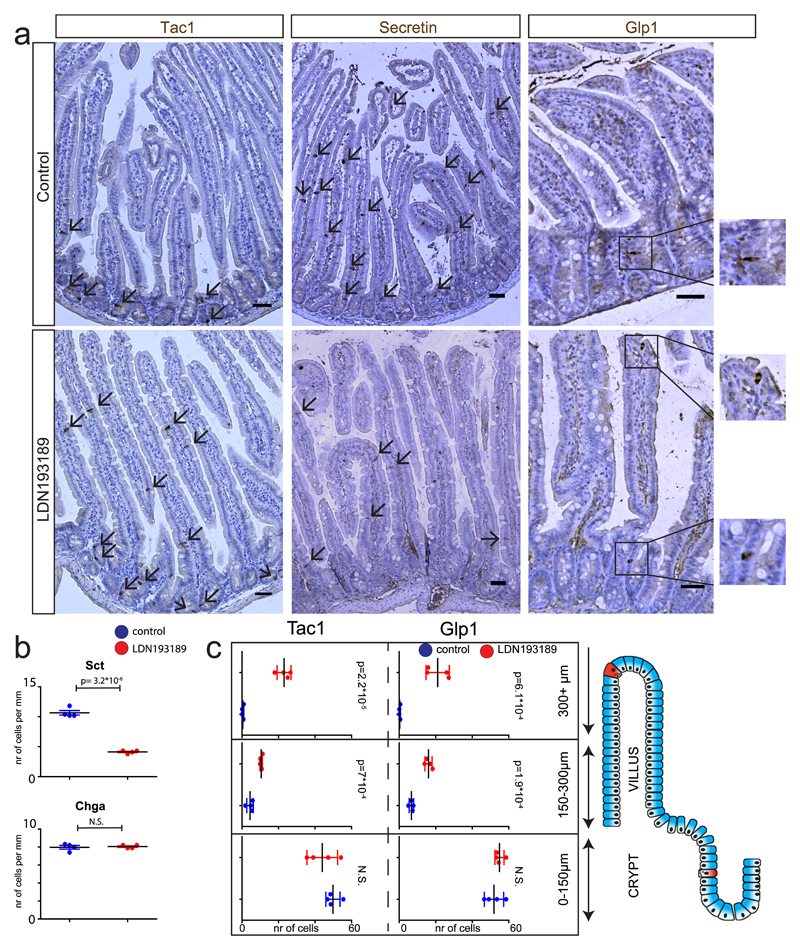
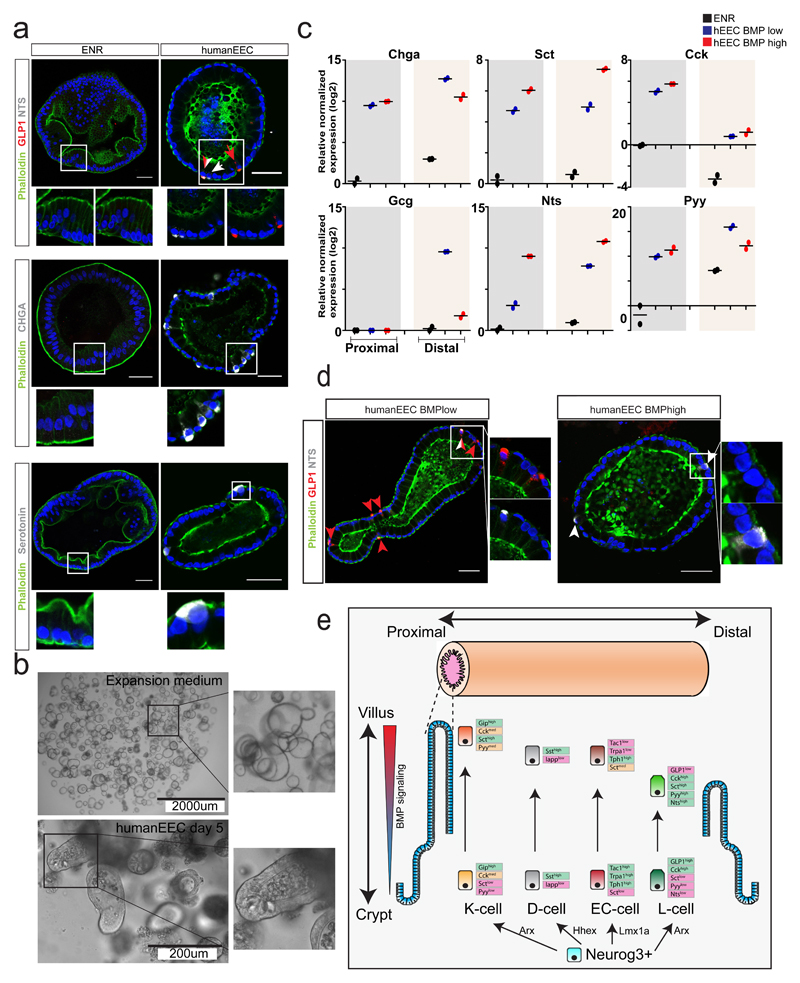
Enteroendocrine cells (EECs) control a wide range of physiological processes linked to metabolism1. We show that EEC hormones are differentially expressed between crypts (for example, Glp1) and villi (for example, secretin). As demonstrated by single-cell mRNA sequencing using murine Lgr5+ cell-derived organoids, BMP4 signals alter the hormone expression profiles of individual EECs to resemble those found in the villus. Accordingly, BMP4 induces hormone switching of EECs migrating up the crypt-villus axis in vivo. Our findings imply that EEC lineages in the small intestine exhibit a more flexible hormone repertoire than previously proposed. We also describe a protocol to generate human EECs in organoids and demonstrate a similar regulation of hormone expression by BMP signalling. These findings establish alternative strategies to target EECs with therapeutically relevant hormone production through BMP modulation.
Conflict of interest statement
The authors disclose no conflicts of interest.
Figures
Comment in
-
Limited gut cell repertoire for multiple hormones.Nat Cell Biol. 2018 Aug;20(8):865-867. doi: 10.1038/s41556-018-0153-9. Nat Cell Biol. 2018. PMID: 30038256 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
