

Parallel circuits from the bed nuclei of stria terminalis to the lateral hypothalamus drive opposing emotional states
- PMID: 30038273
- PMCID: PMC6095688
- DOI: 10.1038/s41593-018-0198-x
Parallel circuits from the bed nuclei of stria terminalis to the lateral hypothalamus drive opposing emotional states
Abstract
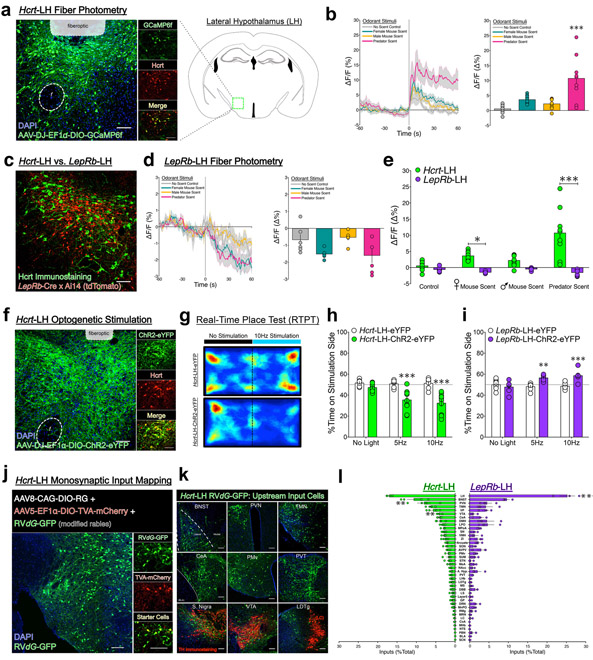
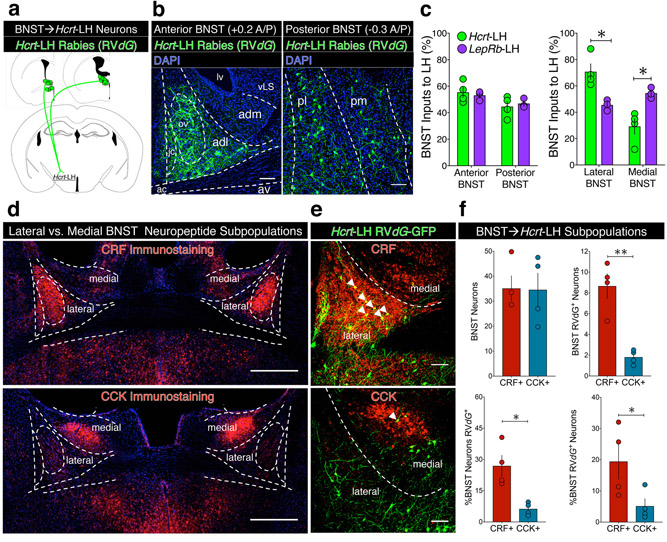
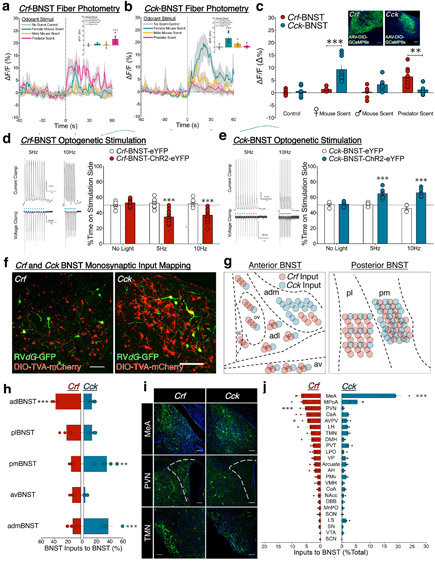
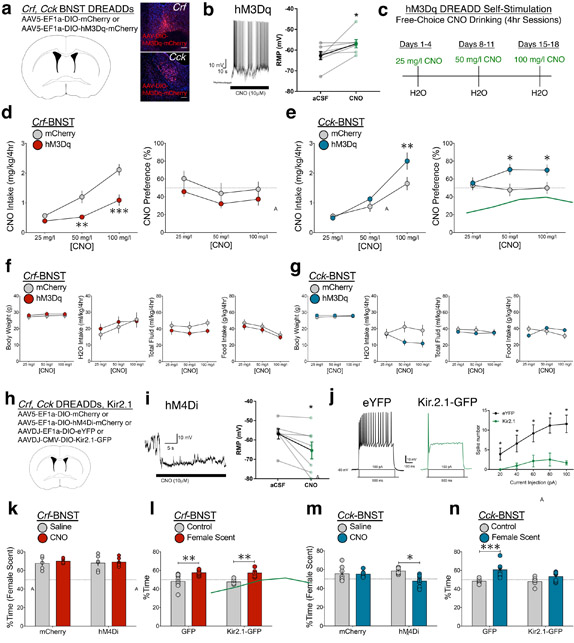
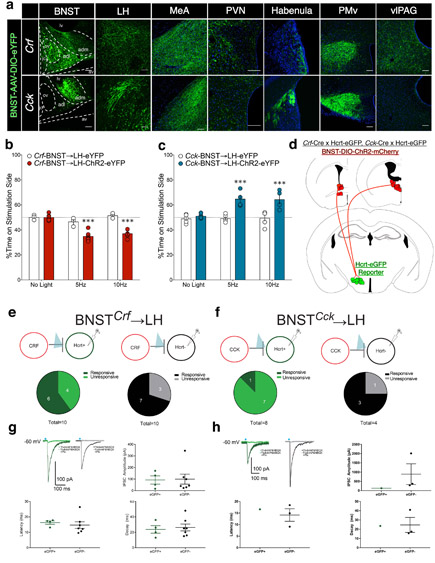
Lateral hypothalamus (LH) neurons containing the neuropeptide hypocretin (HCRT; orexin) modulate affective components of arousal, but their relevant synaptic inputs remain poorly defined. Here we identified inputs onto LH neurons that originate from neuronal populations in the bed nuclei of stria terminalis (BNST; a heterogeneous region of extended amygdala). We characterized two non-overlapping LH-projecting GABAergic BNST subpopulations that express distinct neuropeptides (corticotropin-releasing factor, CRF, and cholecystokinin, CCK). To functionally interrogate BNST→LH circuitry, we used tools for monitoring and manipulating neural activity with cell-type-specific resolution in freely behaving mice. We found that Crf-BNST and Cck-BNST neurons respectively provide abundant and sparse inputs onto Hcrt-LH neurons, display discrete physiological responses to salient stimuli, drive opposite emotionally valenced behaviors, and receive different proportions of inputs from upstream networks. Together, our data provide an advanced model for how parallel BNST→LH pathways promote divergent emotional states via connectivity patterns of genetically defined, circuit-specific neuronal subpopulations.
Figures
References
-
- Sakurai T, et al. Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 92, 573–585 (1998). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
