

Etomoxir Inhibits Macrophage Polarization by Disrupting CoA Homeostasis
- PMID: 30043752
- PMCID: PMC6125190
- DOI: 10.1016/j.cmet.2018.06.001
Etomoxir Inhibits Macrophage Polarization by Disrupting CoA Homeostasis
Abstract
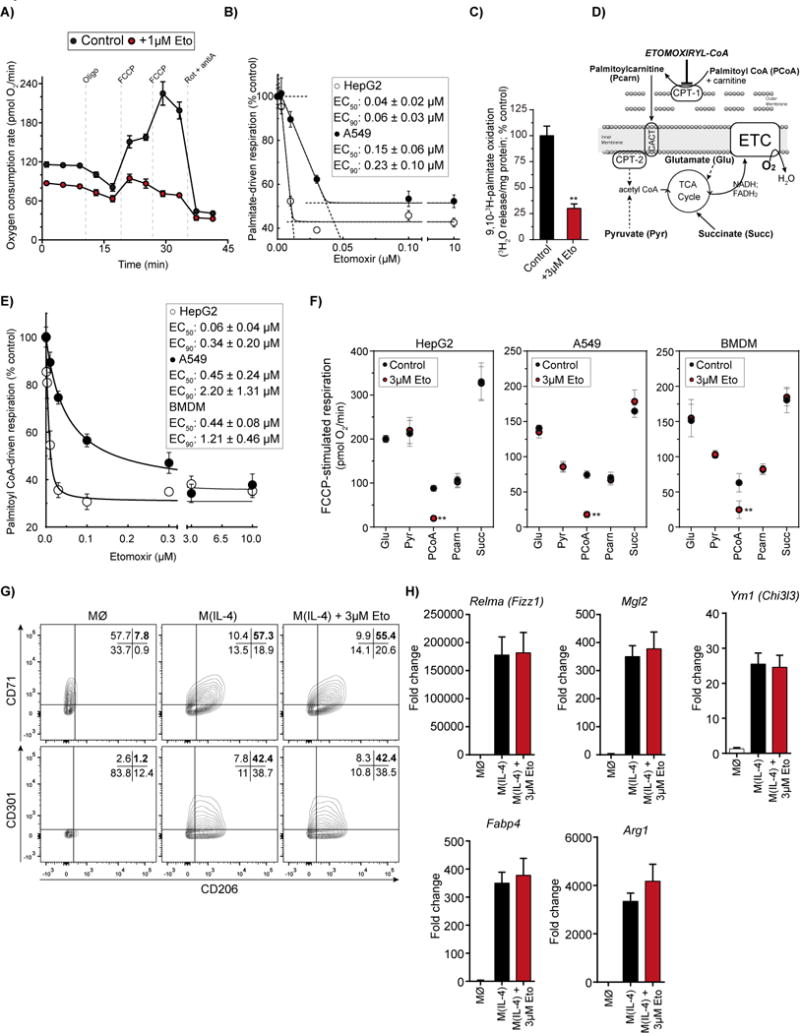
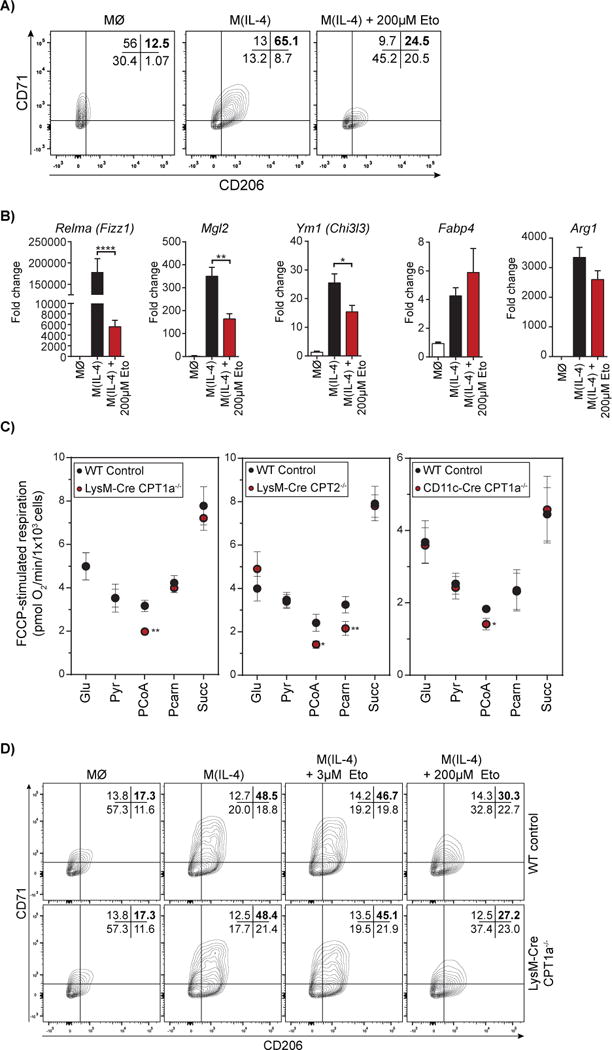
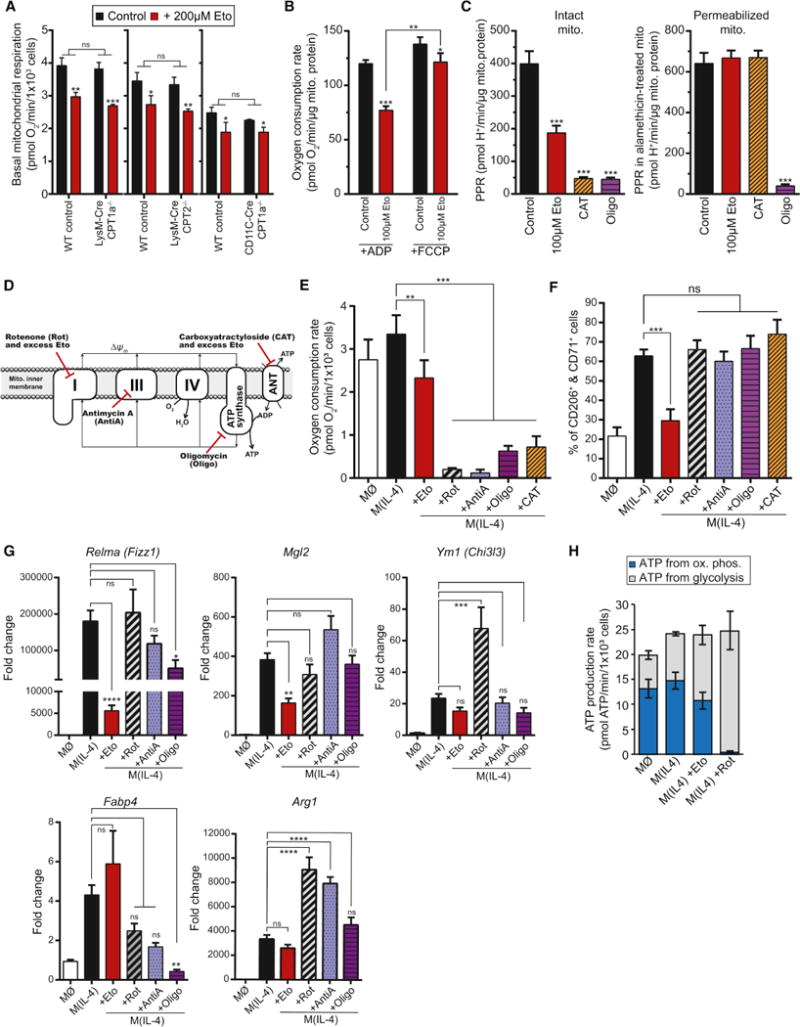
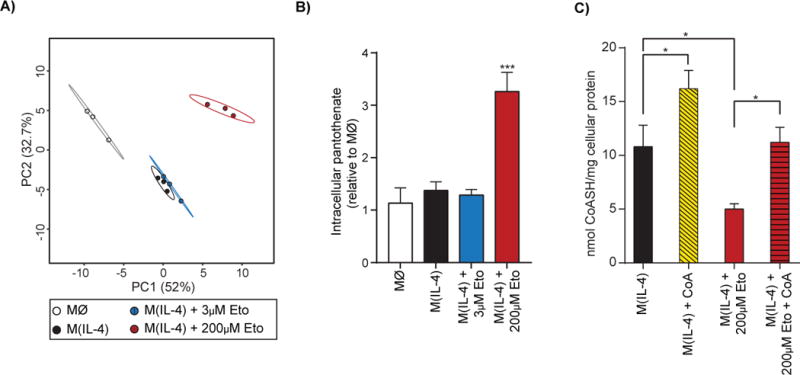
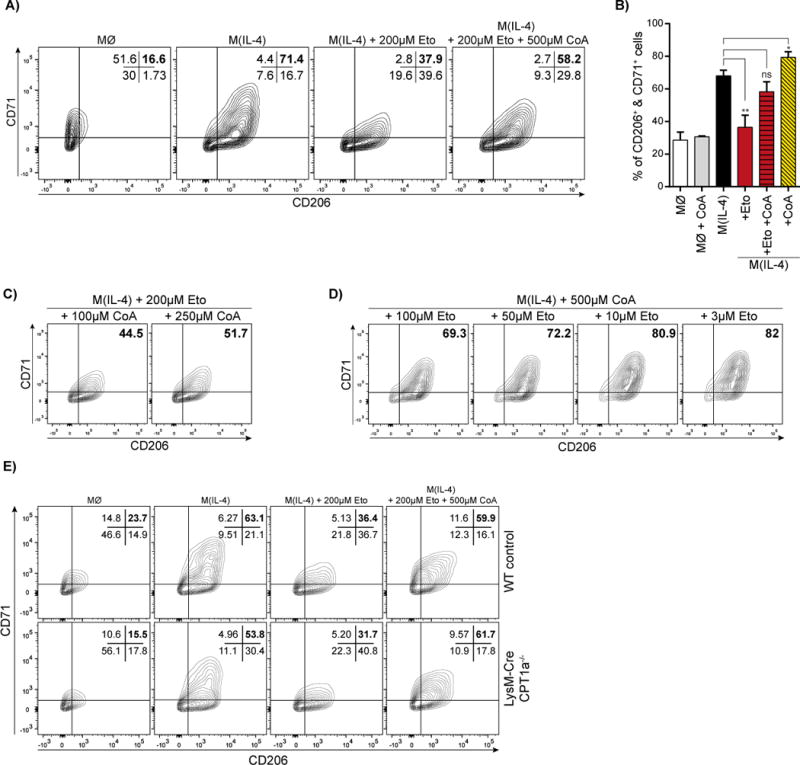
Long-chain fatty acid (LCFA) oxidation has been shown to play an important role in interleukin-4 (IL-4)-mediated macrophage polarization (M(IL-4)). However, many of these conclusions are based on the inhibition of carnitine palmitoyltransferase-1 with high concentrations of etomoxir that far exceed what is required to inhibit enzyme activity (EC90 < 3 μM). We employ genetic and pharmacologic models to demonstrate that LCFA oxidation is largely dispensable for IL-4-driven polarization. Unexpectedly, high concentrations of etomoxir retained the ability to disrupt M(IL-4) polarization in the absence of Cpt1a or Cpt2 expression. Although excess etomoxir inhibits the adenine nucleotide translocase, oxidative phosphorylation is surprisingly dispensable for M(IL-4). Instead, the block in polarization was traced to depletion of intracellular free coenzyme A (CoA), likely resulting from conversion of the pro-drug etomoxir into active etomoxiryl CoA. These studies help explain the effect(s) of excess etomoxir on immune cells and reveal an unappreciated role for CoA metabolism in macrophage polarization.
Keywords: CPT-1; CPT-2; coenzyme A; interleukin 4; long-chain fatty acid oxidation; macrophage polarization; mitochondria; oxidative phosphorylation; pantothenate.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
K.C., B.P.D., D.A.F., and G.W.R. are employees and shareholders of Agilent Technologies.
Figures
Comment in
-
Fatty Acid Oxidation in Macrophages and T Cells: Time for Reassessment?Cell Metab. 2018 Oct 2;28(4):538-540. doi: 10.1016/j.cmet.2018.09.018. Cell Metab. 2018. PMID: 30282046
References
-
- Agius L, Meredith EJ, Sherratt H, Stanley A. Stereospecificity of the inhibition by etomoxir of fatty acid and cholesterol synthesis in isolated rat hepatocytes. Biochem Pharmacol. 1991;42:1717–1720. - PubMed
-
- Bartlett K, Bone AJ, Koundakjian P, Meredith E, Turnbull DM, Sherratt HS. Inhibition of mitochondrial β-oxidation at the stage of carnitine palmitoyltransferase I by the coenzyme A esters of some substituted hypoglycaemic oxirane-carboxylic acids. Biochem Soc Trans. 1981;9:574–575.
-
- Bentebibel A, Sebastian D, Herrero L, Lopez-Vinas E, Serra D, Asins G, Gomez-Puertas P, Hegardt FG. Novel effect of C75 on carnitine palmitoyltransferase I activity and palmitate oxidation. Biochemistry. 2006;45:4339–4350. - PubMed
-
- Ceccarelli SM, Chomienne O, Gubler M, Arduini A. Carnitine Palmitoyltransferase (CPT) Modulators: A Medicinal Chemistry Perspective on 35 Years of Research. J Med Chem. 2011;54:3109–3152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
