

M current regulates firing mode and spike reliability in a collision-detecting neuron
- PMID: 30044671
- PMCID: PMC6230786
- DOI: 10.1152/jn.00363.2018
M current regulates firing mode and spike reliability in a collision-detecting neuron
Abstract
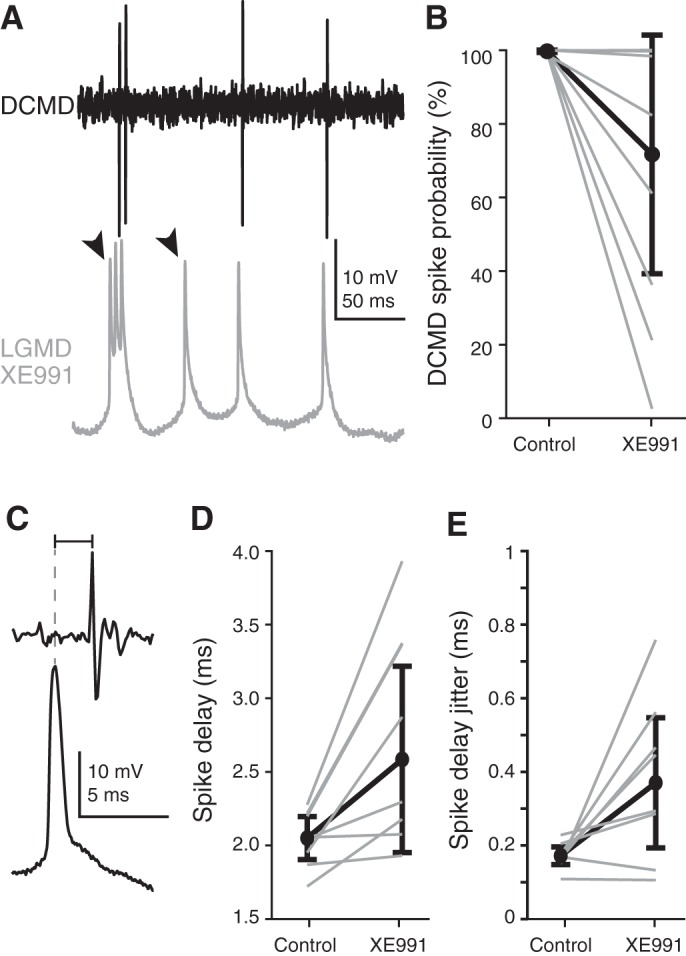
All animals must detect impending collisions to escape and reliably discriminate them from nonthreatening stimuli, thus preventing false alarms. Therefore, it is no surprise that animals have evolved highly selective and sensitive neurons dedicated to such tasks. We examined a well-studied collision-detection neuron in the grasshopper ( Schistocerca americana) using in vivo electrophysiology, pharmacology, and computational modeling. This lobula giant movement detector (LGMD) neuron is excitable by inputs originating from each ommatidia of the compound eye. It possesses many intrinsic properties that increase its selectivity to objects approaching on a collision course, including switching between burst and nonburst firing. In this study, we demonstrate that the LGMD neuron exhibits a large M current, generated by noninactivating K+ channels, that shortens the temporal window of dendritic integration, regulates a firing mode switch between burst and isolated spiking, increases the precision of spike timing, and increases the reliability of spike propagation to downstream motor centers. By revealing how the M current increases the LGMD's ability to detect impending collisions, our results suggest that similar channels may play an analogous role in other collision detection circuits. NEW & NOTEWORTHY The ability to reliably detect impending collisions is a critical survival skill. The nervous systems of many animals have developed dedicated neurons for accomplishing this task. We used a mix of in vivo electrophysiology and computational modeling to investigate the role of M potassium channels within one such collision-detecting neuron and show that through regulation of burst firing and enhancement of spiking reliability, the M current increases the ability to detect impending collisions.
Keywords: M current; burst firing; collision avoidance; lobula giant movement detector; spike timing.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases