

Schistosoma japonicum IAP and Teg20 safeguard tegumental integrity by inhibiting cellular apoptosis
- PMID: 30044778
- PMCID: PMC6078320
- DOI: 10.1371/journal.pntd.0006654
Schistosoma japonicum IAP and Teg20 safeguard tegumental integrity by inhibiting cellular apoptosis
Abstract
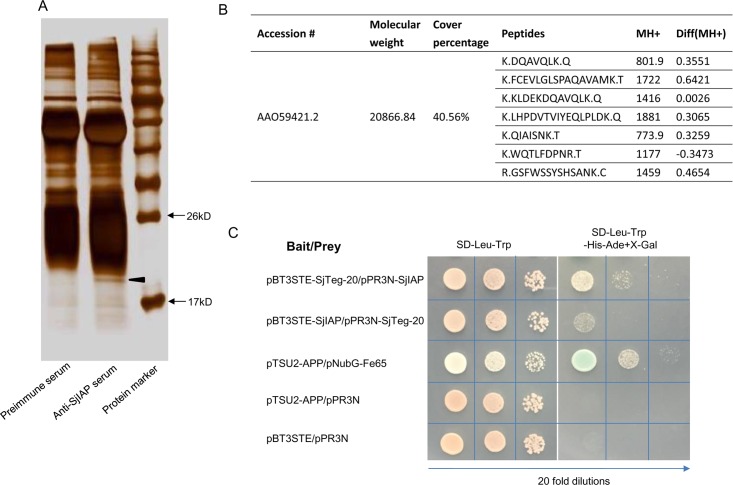
Schistosomes are causative agents of human schistosomiasis, which is endemic in tropical and subtropical areas of the world. Adult schistosomes can survive in their final hosts for several decades, and they have evolved various strategies to overcome the host immune response. Consequently, understanding the mechanisms that regulate parasitic cell survival will open avenues for developing novel strategies against schistosomiasis. Our previous study suggested that an inhibitor of apoptosis protein in Schistosoma japonicum (SjIAP) may play important roles in parasitic survival and development. Here, we demonstrated that SjIAP can negatively regulate cellular apoptosis in S. japonicum by suppressing caspase activity. Immunohistochemistry analysis indicated that SjIAP ubiquitously expressed within the worm body including the tegument. Silencing of SjIAP expression via small interfering RNA led to destruction of the tegument integrity in schistosomes. We further used co-immunoprecipitation to identify interaction partners of SjIAP and revealed the tegument protein SjTeg-20 as a putative interacting partner of SjIAP. The interaction between SjIAP and SjTeg-20 was confirmed by a yeast two-hybrid (Y2H) assay. Moreover, results of a TUNEL assay, RNA interference, scanning and transmission electron microscopy, caspase assays, transcript profiling, and protein localization of both interacting molecules provided first evidence for an essential role of SjIAP and SjTeg-20 to maintain the structural integrity of the tegument by negatively regulating apoptosis. Taken together, our findings suggest that the cooperative activities of SjIAP and SjTeg-20 belong to the strategic inventory of S. japonicum ensuring survival in the hostile environment within the vasculature of the final host.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
-
- Williams GT. Programmed cell death: a fundamental protective response to pathogens. Trends Microbiol. 1994;2(12):463–4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
