

Schlafen 12 Interaction with SerpinB12 and Deubiquitylases Drives Human Enterocyte Differentiation
- PMID: 30045019
- PMCID: PMC6123821
- DOI: 10.1159/000492019
Schlafen 12 Interaction with SerpinB12 and Deubiquitylases Drives Human Enterocyte Differentiation
Abstract
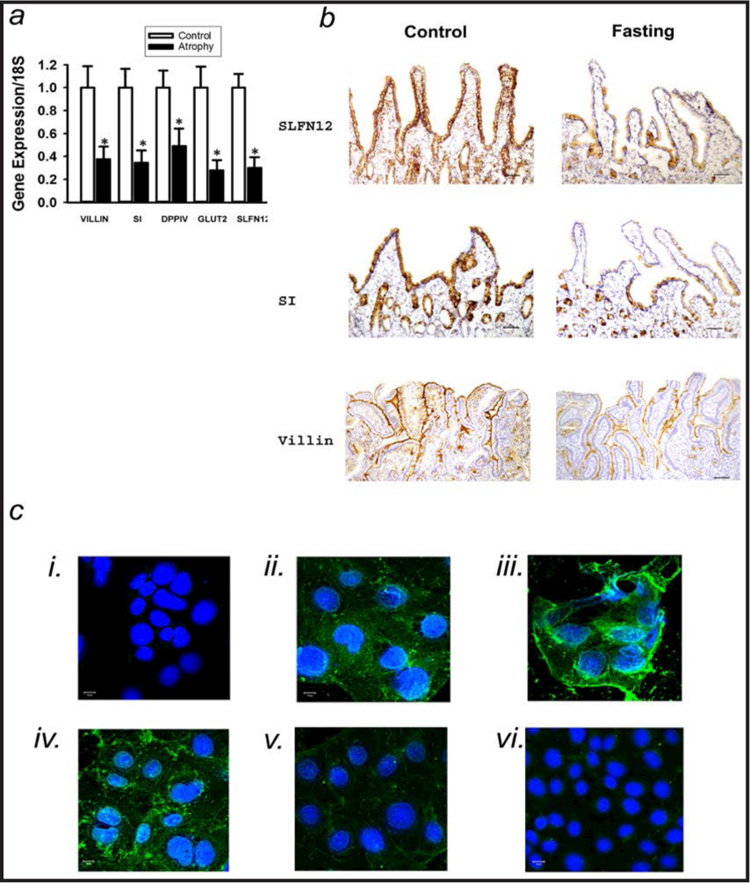
Background/aims: Human enterocytic differentiation is altered during development, fasting, adaptation, and bariatric surgery, but its intracellular control remains unclear. We hypothesized that Schlafen 12 (SLFN12) regulates enterocyte differentiation.
Methods: We used laser capture dissection of epithelium, qRT-PCR, and immunohistochemistry to evaluate SLFN12 expression in biopsies of control and fasting human duodenal mucosa, and viral overexpression and siRNA to trace the SLFN12 pathway in human Caco-2 and HIEC6 intestinal epithelial cells.
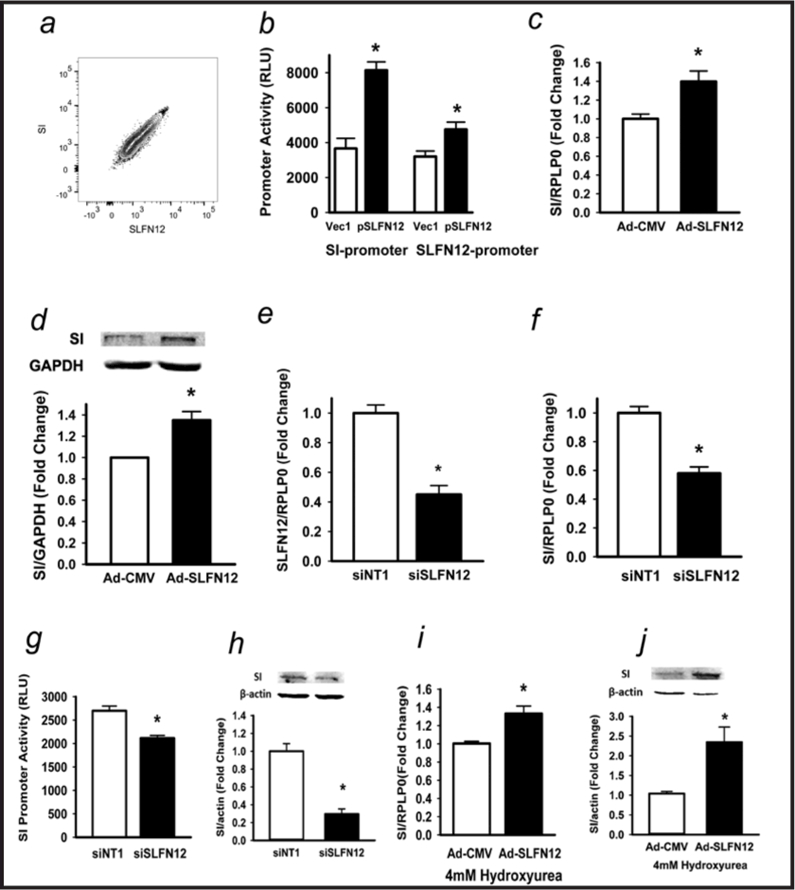
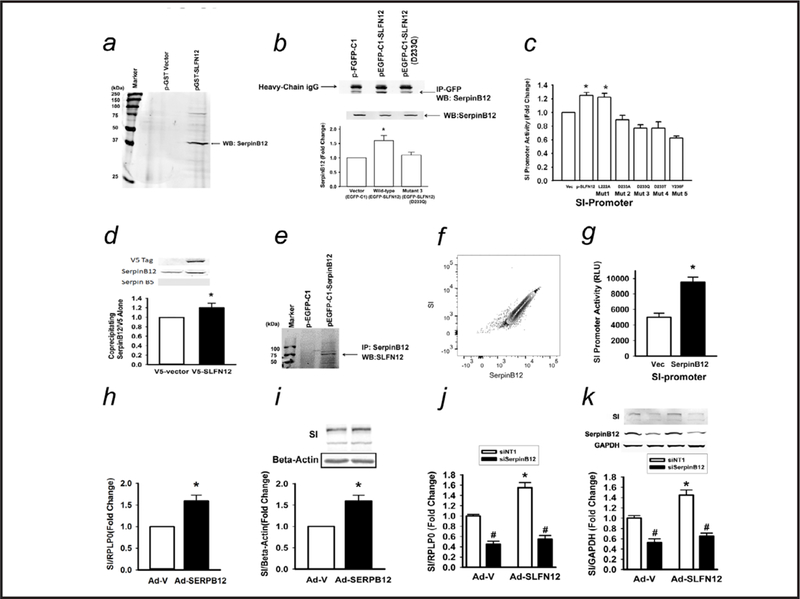
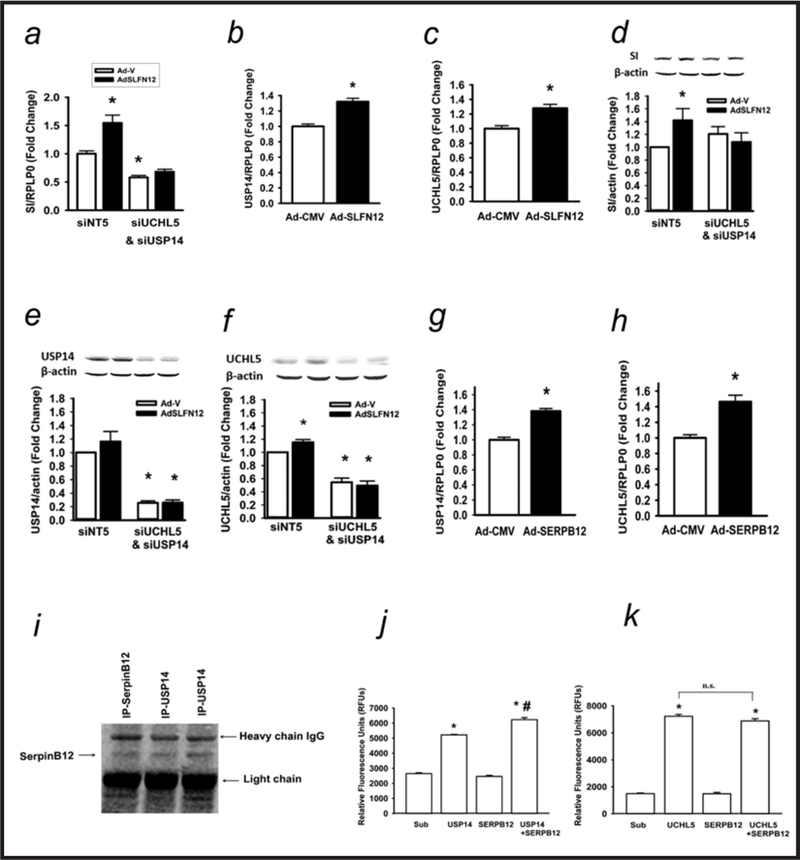
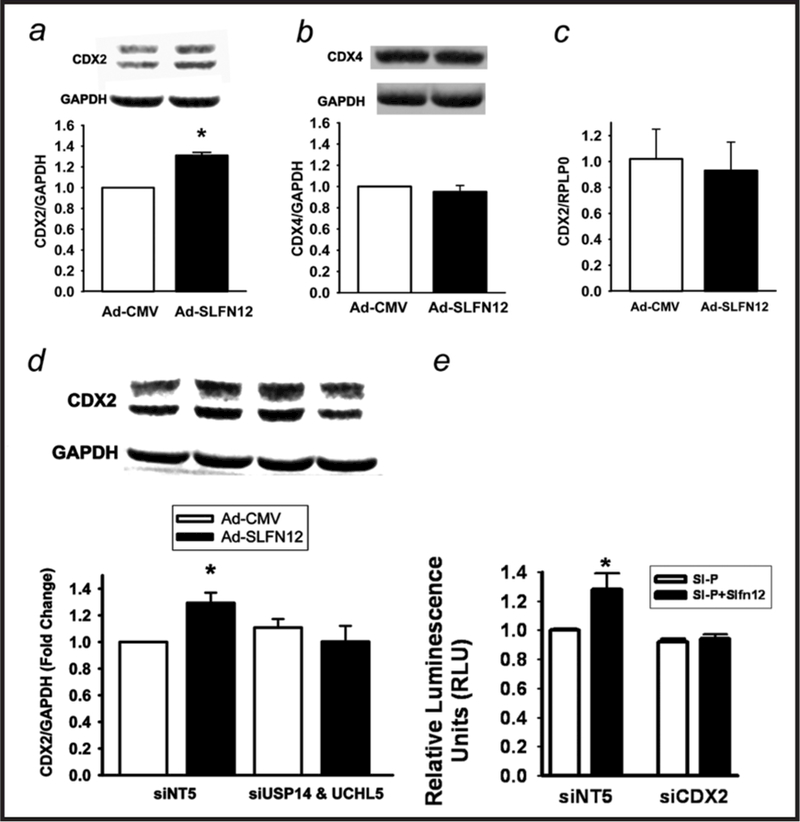
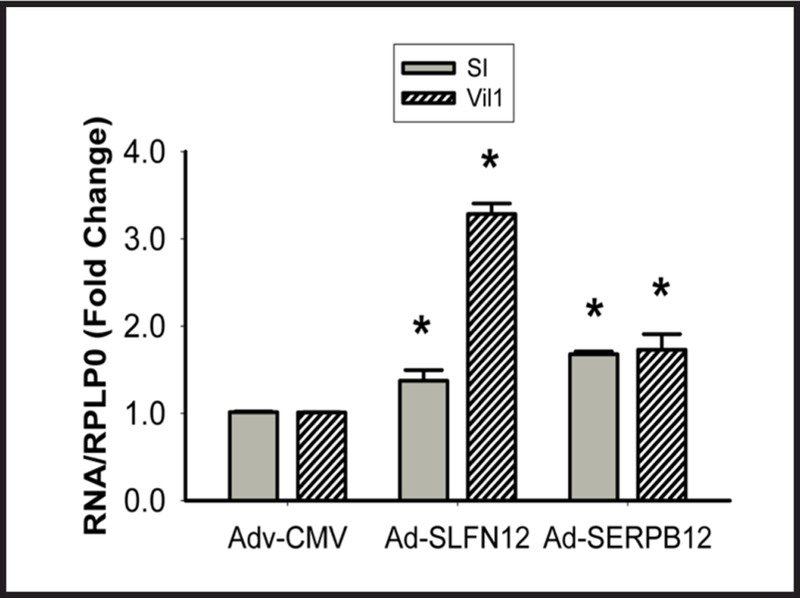
Results: Fasting human duodenal mucosa expressed less SLFN12 mRNA and protein, accompanied by decreases in enterocytic markers like sucrase-isomaltase. SLFN12 overexpression increased Caco-2 sucrase-isomaltase promoter activity, mRNA, and protein independently of proliferation, and activated the SLFN12 putative promoter. SLFN12 coprecipitated Serpin B12 (SERPB12). An inactivating SLFN12 point mutation prevented both SERPB12 binding and sucrase-isomaltase induction. SERPB12 overexpression also induced sucrase-isomaltase, while reducing SERPB12 prevented the SLFN12 effect on sucrase-isomaltase. Sucrase-isomaltase induction by both SLFN12 and SERPB12 was attenuated by reducing UCHL5 or USP14, and blocked by reducing both. SERPB12 stimulated USP14 but not UCHL5 activity. SERPB12 coprecipitated USP14 but not UCHL5. Moreover, SLFN12 increased protein levels of the sucrase-isomaltase-promoter-binding transcription factor cdx2 without altering Cdx2 mRNA. This was prevented by reducing UCHL5 and USP14. We further validated this pathway in vitro and in vivo. SLFN12 or SERPB12 overexpression induced sucrase-isomaltase in human non-malignant HIEC-6 enterocytes.
Conclusions: SLFN12 regulates human enterocytic differentiation by a pathway involving SERPB12, the deubiquitylases, and Cdx2. This pathway may be targeted to manipulate human enterocytic differentiation in mucosal atrophy, short gut or obesity.
Keywords: Differentiation; Epithelium; Intestine; Schlafen 12; Serpin B12; Signaling; UCHL5; USP14.
© 2018 The Author(s). Published by S. Karger AG, Basel.
Conflict of interest statement
Disclosure Statement
The authors declare that they have no competing interests.
Figures
References
-
- Wales PW, Christison-Lagay ER: Short bowel syndrome: epidemiology and etiology. Semin Pediatr Surg 2010;19:3–9. - PubMed
-
- Basson MD: In vitro evidence for matrix regulation of intestinal epithelial biology during mucosal healing. Life Sci 2001;69:3005–3018. - PubMed
-
- Basson MD, Li GD, Hong F, Han O, Sumpio BE: Amplitude-dependent modulation of brush border enzymes and proliferation by cyclic strain in human intestinal Caco-2 monolayers. J Cell Physiol 1996;168:476–488. - PubMed
-
- Emenaker NJ, Basson MD: Short chain fatty acids inhibit human (SW1116) colon cancer cell invasion by reducing urokinase plasminogen activator activity and stimulating TIMP-1 and TIMP-2 activities, rather than via MMP modulation. J Surg Res 1998;76:41–46. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials