

Delimiting the autoinhibitory module of von Willebrand factor
- PMID: 30053340
- PMCID: PMC6173635
- DOI: 10.1111/jth.14251
Delimiting the autoinhibitory module of von Willebrand factor
Abstract
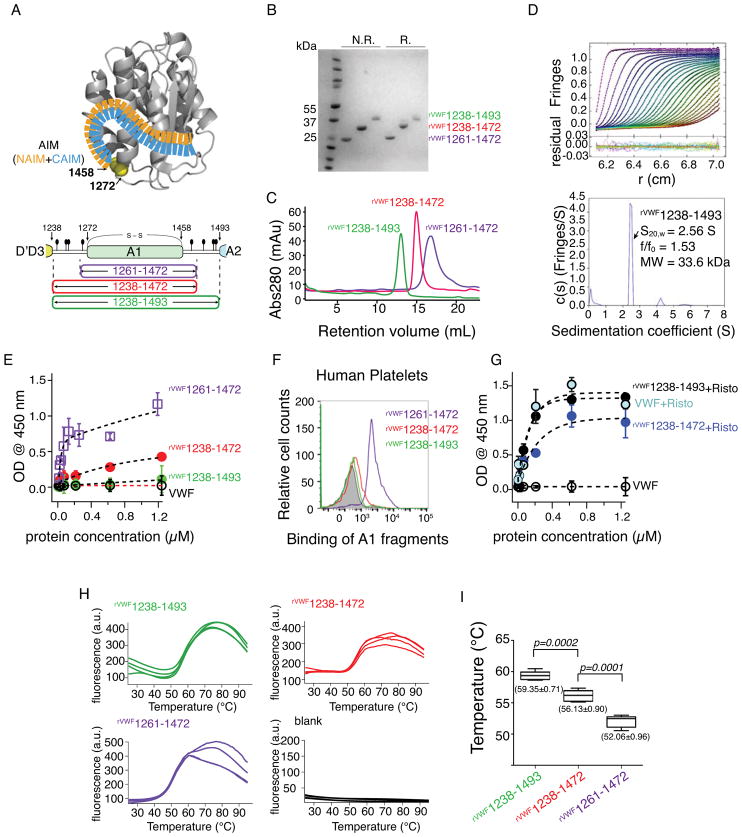
Essentials The self-inhibitory mechanism of von Willebrand factor (VWF) remains unclear. Residues flanking the A1 domain of VWF form a discontinuous autoinhibitory module (AIM). rVWF1238-1493 exhibited greater thermostability and inactivity than its shorter counterparts. The cooperative coupling between the N- and C- AIM regions are required for inhibiting A1.
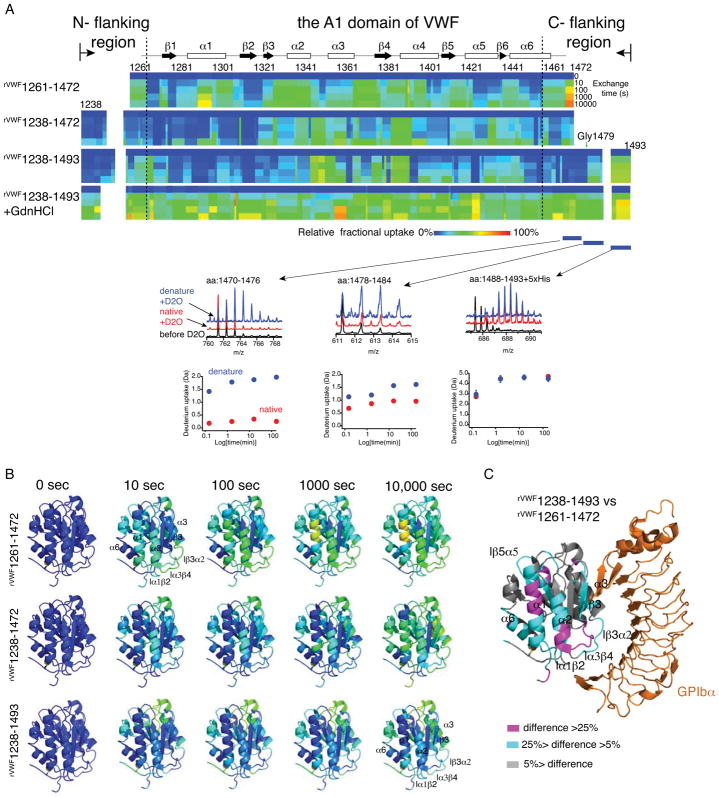
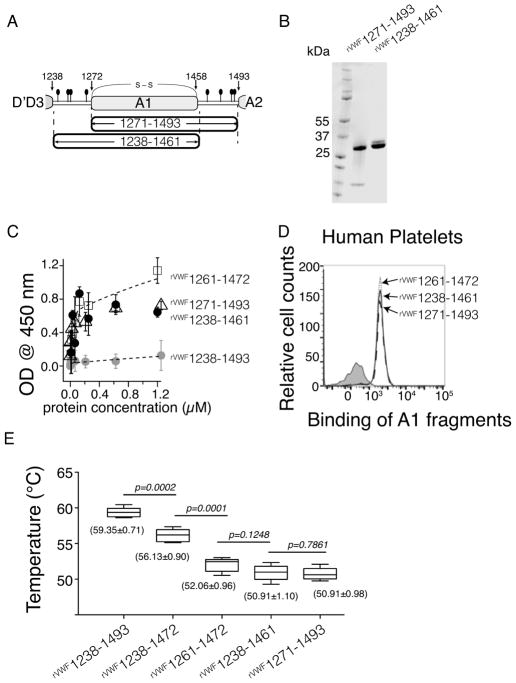
Summary: Background The hierarchical hemostasis response involves a self-inhibitory feature of von Willebrand factor (VWF) that has not been fully characterized. The residues flanking the A1 domain of VWF are important in this self-inhibition by forming an autoinhibitory module (AIM) that masks the A1 domain. Objectives To delimit the AIM sequence and to evaluate the cooperative interplay between the discontinuous AIM regions. Methods ELISA, flow cytometry, a thermal stability assay and hydrogen-deuterium exchange (HDX) mass spectrometry were used to characterize recombinant VWF A1 fragments varying in length. Results The longest A1 fragment (rVWF1238-1493 ) showed higher inactivity in binding the platelet receptor glycoprotein (GP) Ibα and greater thermostability than its shorter counterparts. The HDX results showed that most of the N-terminal residues and residues 1459-1478 at the C-terminus of rVWF1238-1493 have slower deuterium uptake than the residues in its denatured counterpart, implying that these residues may interact with the A1 domain. In contrast, residues 1479-1493 showed less difference from the denatured form, indicating that these residues are unlikely to be involved in binding the A1 domain. The A1 fragment that lacks either the entire C-terminal flanking region of the AIM (C-AIM), i.e. rVWF1238-1461 , or the entire N-terminal flanking region of the AIM (N-AIM), i.e. rVWF1271-1493 , showed high GPIbα-binding affinity and low thermostability, suggesting that removal of either N-terminal or C-terminal residues resulted in loss of AIM inhibition of the A1 domain. Conclusion The AIM is probably composed of residues 1238-1271 (N-AIM) and 1459-1478 (C-AIM). Neither the N-AIM nor the C-AIM alone could fully inhibit binding of the A1 domain to GPIbα.
Keywords: blood platelets; hemostasis; platelet GPIb-IX complex; tandem mass spectrometry; von Willebrand factor.
© 2018 International Society on Thrombosis and Haemostasis.
Conflict of interest statement
The authors state that they have no conflict of interest.
Figures
References
-
- Slayter H, Loscalzo J, Bockenstedt P, Handin RI. Native conformation of human von Willebrand protein. Analysis by electron microscopy and quasi-elastic light scattering. The Journal of biological chemistry. 1985;260:8559–63. - PubMed
-
- Cruz MA, Handin RI, Wise RJ. The interaction of the von Willebrand factor-A1 domain with platelet glycoprotein Ib/IX. The role of glycosylation and disulfide bonding in a monomeric recombinant A1 domain protein. J Biol Chem. 1993;268:21238–45. - PubMed
-
- Miyata S, Goto S, Federici AB, Ware J, Ruggeri ZM. Conformational changes in the A1 domain of von Willebrand factor modulating the interaction with platelet glycoprotein Ibalpha. The Journal of biological chemistry. 1996;271:9046–53. - PubMed
-
- Miura S, Fujimura Y, Sugimoto M, Kawasaki T, Ikeda Y, Titani K, Yoshioka A. Structural elements influencing von Willebrand factor (vWF) binding affinity for platelet glycoprotein Ib within a dispase-digested vWF fragment. Blood. 1994;84:1553–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
