

iac Gene Expression in the Indole-3-Acetic Acid-Degrading Soil Bacterium Enterobacter soli LF7
- PMID: 30054366
- PMCID: PMC6146976
- DOI: 10.1128/AEM.01057-18
iac Gene Expression in the Indole-3-Acetic Acid-Degrading Soil Bacterium Enterobacter soli LF7
Abstract
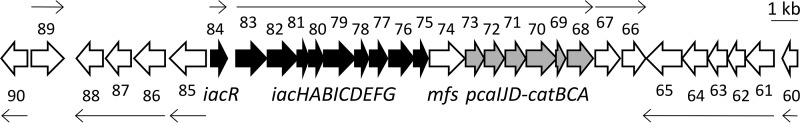
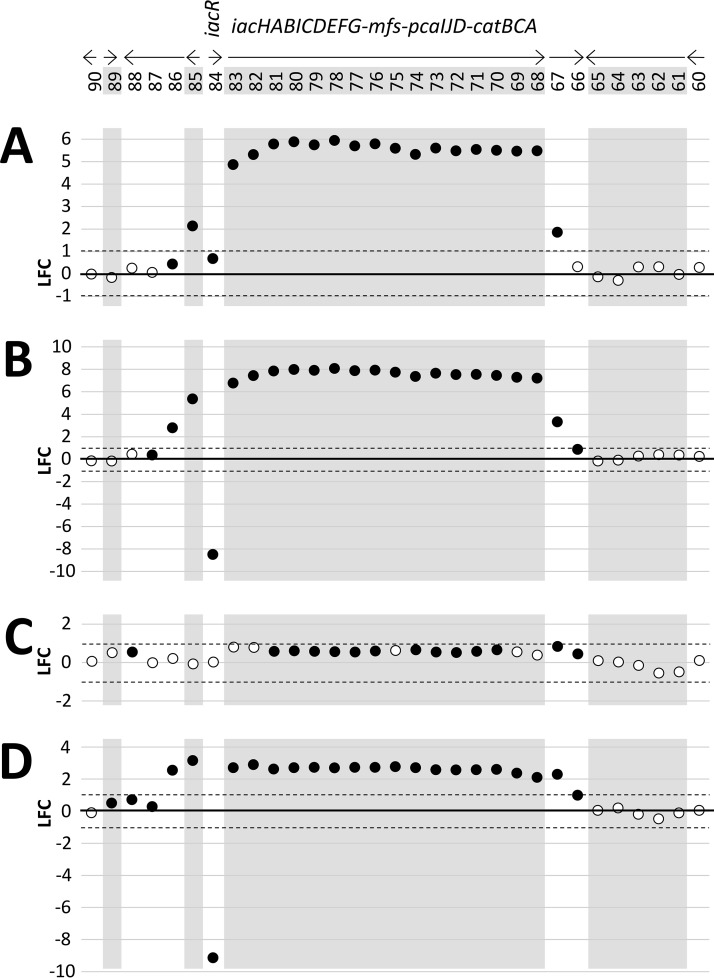
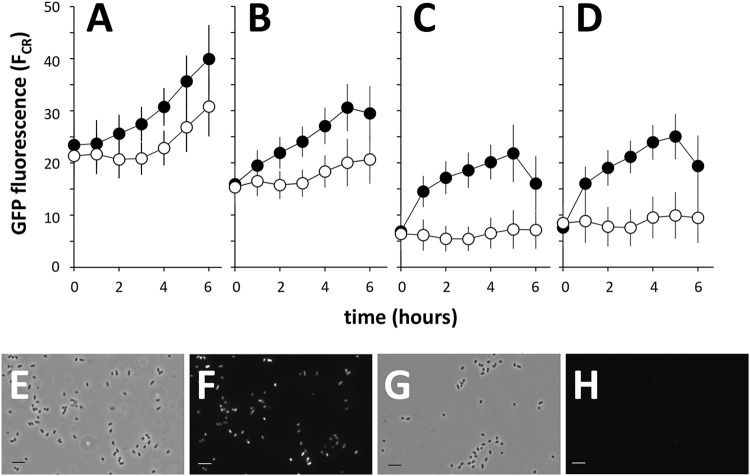
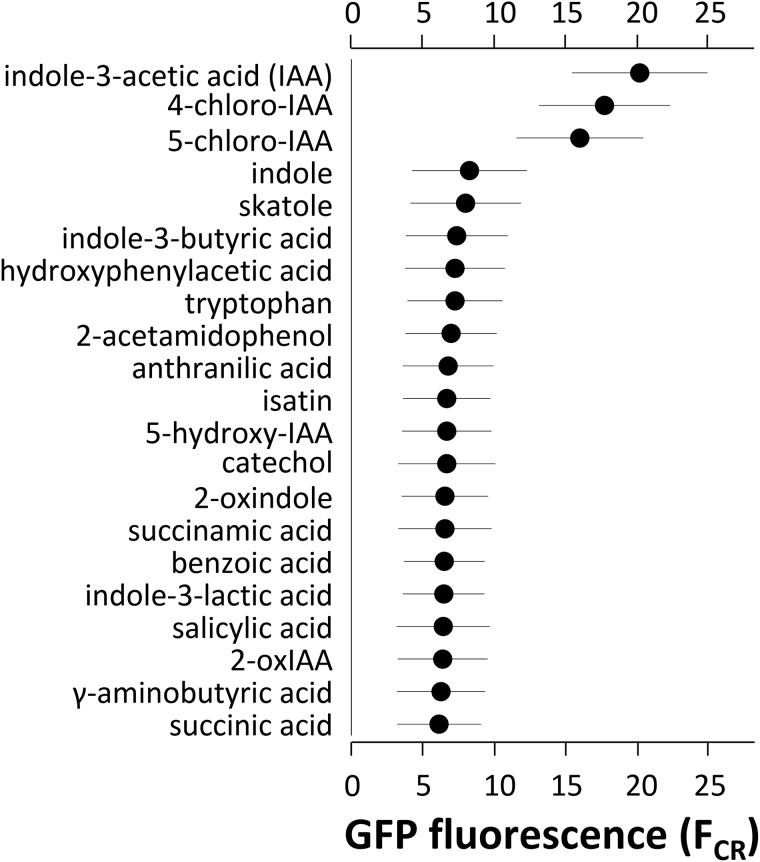
We show for soil bacterium Enterobacter soli LF7 that the possession of an
Keywords: Enterobacter asburiae LF7a; Enterobacter soli LF7; IAA; auxin; iac genes; indole-3-acetic acid catabolism.
Copyright © 2018 American Society for Microbiology.
Figures

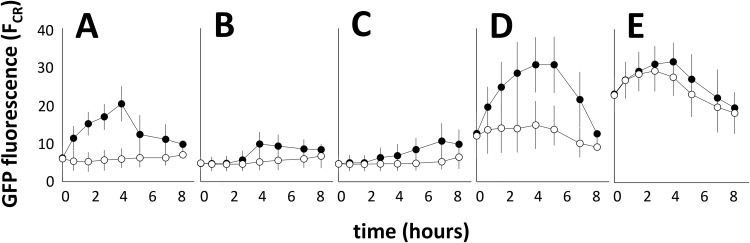
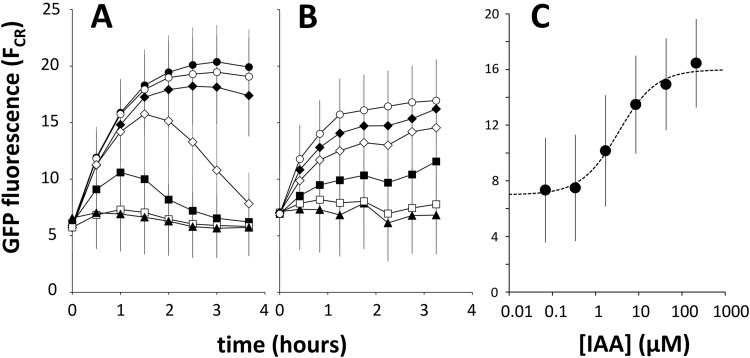
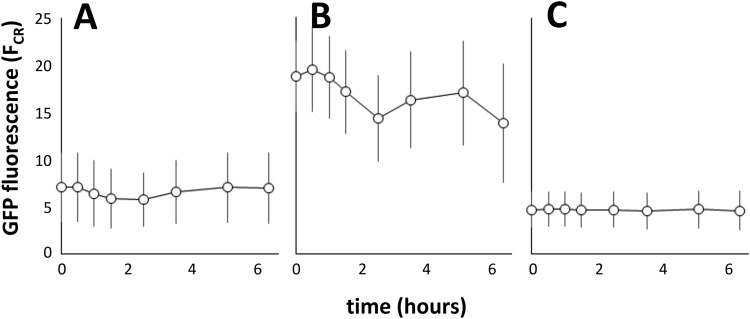
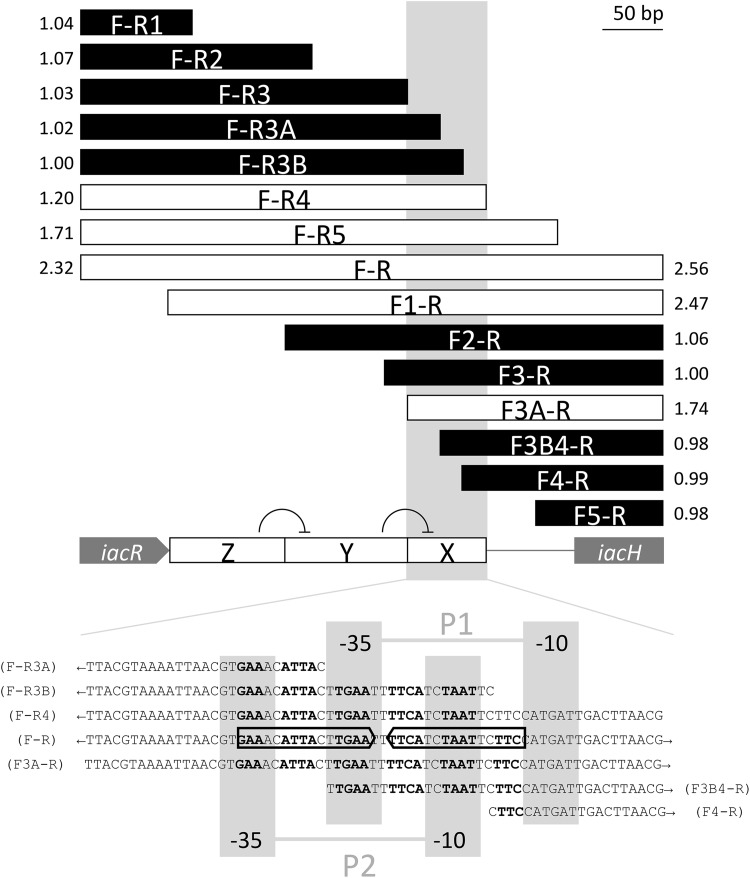
References
-
- Lin GH, Chen HP, Huang JH, Liu TT, Lin TK, Wang SJ, Tseng CH, Shu HY. 2012. Identification and characterization of an indigo-producing oxygenase involved in indole 3-acetic acid utilization by Acinetobacter baumannii. Antonie Van Leeuwenhoek 101:881–890. doi: 10.1007/s10482-012-9704-4. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
