

ClpB mutants of Francisella tularensis subspecies holarctica and tularensis are defective for type VI secretion and intracellular replication
- PMID: 30054549
- PMCID: PMC6063899
- DOI: 10.1038/s41598-018-29745-4
ClpB mutants of Francisella tularensis subspecies holarctica and tularensis are defective for type VI secretion and intracellular replication
Abstract
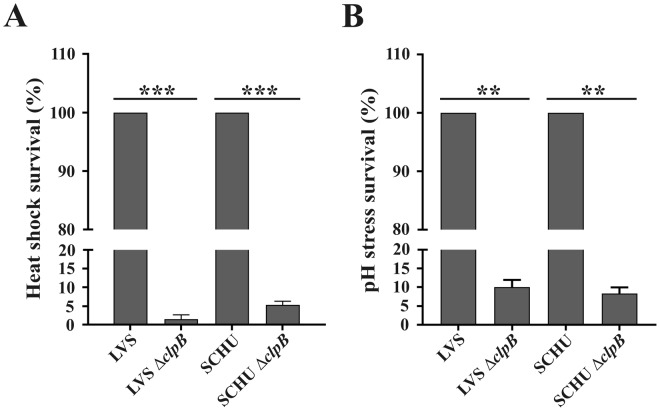
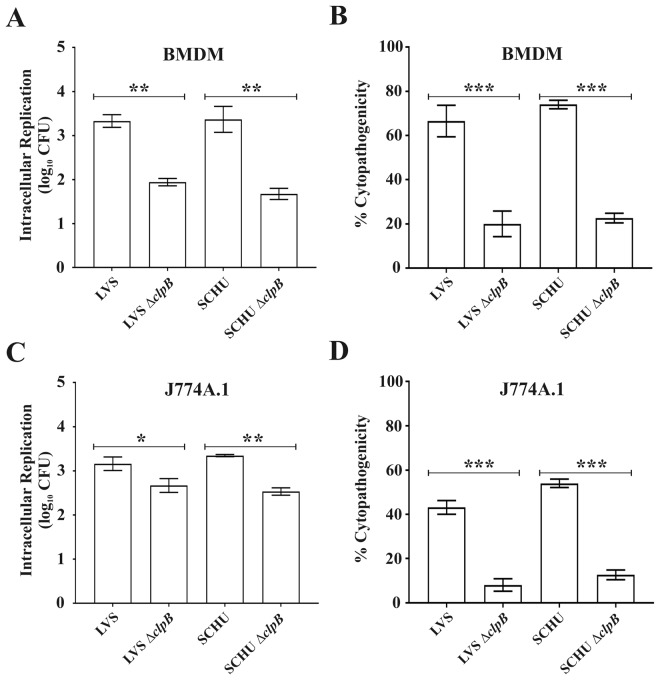
Francisella tularensis, a highly infectious, intracellular bacterium possesses an atypical type VI secretion system (T6SS), which is essential for the virulence of the bacterium. Recent data suggest that the HSP100 family member, ClpB, is involved in T6SS disassembly in the subspecies Francisella novicida. Here, we investigated the role of ClpB for the function of the T6SS and for phenotypic characteristics of the human pathogenic subspecies holarctica and tularensis. The ∆clpB mutants of the human live vaccine strain, LVS, belonging to subspecies holarctica, and the highly virulent SCHU S4 strain, belonging to subspecies tularensis, both showed extreme susceptibility to heat shock and low pH, severely impaired type VI secretion (T6S), and significant, but impaired intracellular replication compared to the wild-type strains. Moreover, they showed essentially intact phagosomal escape. Infection of mice demonstrated that both ΔclpB mutants were highly attenuated, but the SCHU S4 mutant showed more effective replication than the LVS strain. Collectively, our data demonstrate that ClpB performs multiple functions in the F. tularensis subspecies holarctica and tularensis and its function is important for T6S, intracellular replication, and virulence.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- 2013-4581/Vetenskapsrådet (Swedish Research Council)/International
- 2013-8621/Vetenskapsrådet (Swedish Research Council)/International
- 13-4581/Vetenskapsrådet (Swedish Research Council)/International
- 2013-8621/Vetenskapsrådet (Swedish Research Council)/International
LinkOut - more resources
Full Text Sources
Other Literature Sources
