

Review
doi: 10.1038/s41576-018-0034-x.
High-throughput determination of RNA structures
Affiliations
- PMID: 30054568
- PMCID: PMC7388734
- DOI: 10.1038/s41576-018-0034-x
Item in Clipboard
Review
High-throughput determination of RNA structures
Nat Rev Genet.
2018 Oct.
Abstract
RNA performs and regulates a diverse range of cellular processes, with new functional roles being uncovered at a rapid pace. Interest is growing in how these functions are linked to RNA structures that form in the complex cellular environment. A growing suite of technologies that use advances in RNA structural probes, high-throughput sequencing and new computational approaches to interrogate RNA structure at unprecedented throughput are beginning to provide insights into RNA structures at new spatial, temporal and cellular scales.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests.
Figures
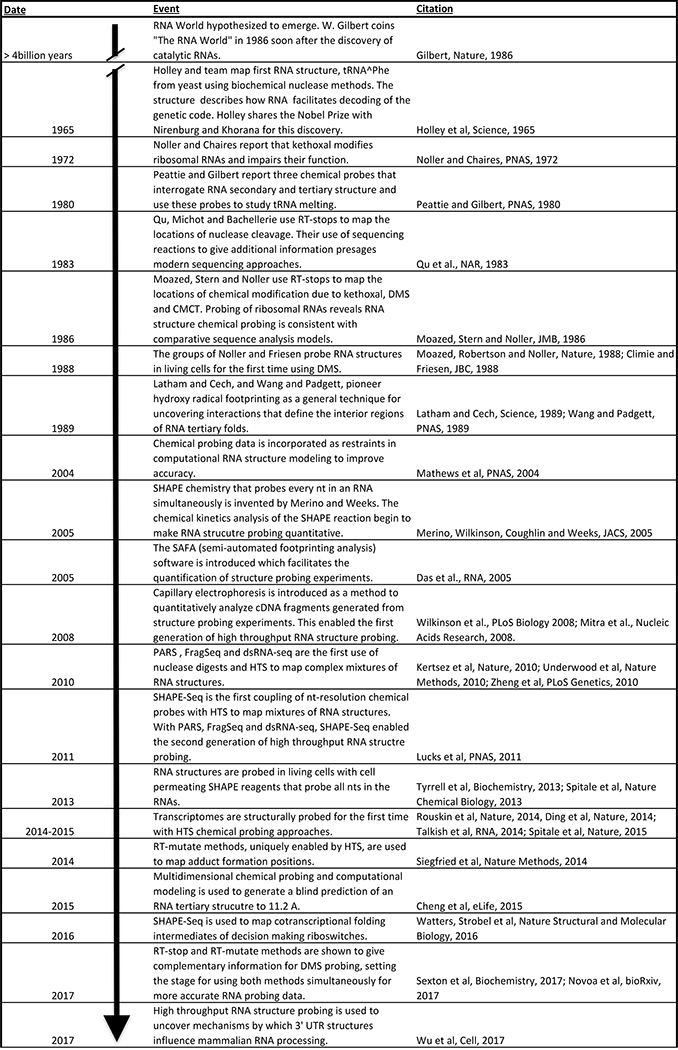
Biochemical probing of RNA structure began even before the diverse and important roles of RNA were fully appreciated. Early methods used sequence and structure dependent cleavage by nuclease enzymes, which gave the first insights into how tRNA structures decode the genetic code. Chemical probes were developed later,,, followed by methods that used RT primer extension to read out adduct position,. Almost two decades later, biochemical probing data was incorporated into RNA folding algorithms to give more accurate models of RNA structure. The development of SHAPE chemistry in 2005 launched efforts to make RNA structure probing quantitative. The coupling of enzymatic,, and chemical probing to high throughput sequencing, and the development of probes that penetrate cell membranes, brought RNA structure probing into the ‘omics’ era. This allowed rapid advances in probing entire transcriptomes–, high resolution modelling of tertiary structures, mapping of nascent RNA cotranscriptional folding pathways, and uncovering the role of RNA structures in mammalian RNA processing. Rapid technological advances such as new adduct mapping methods, promise to continue to increase the accuracy and power of these methods.
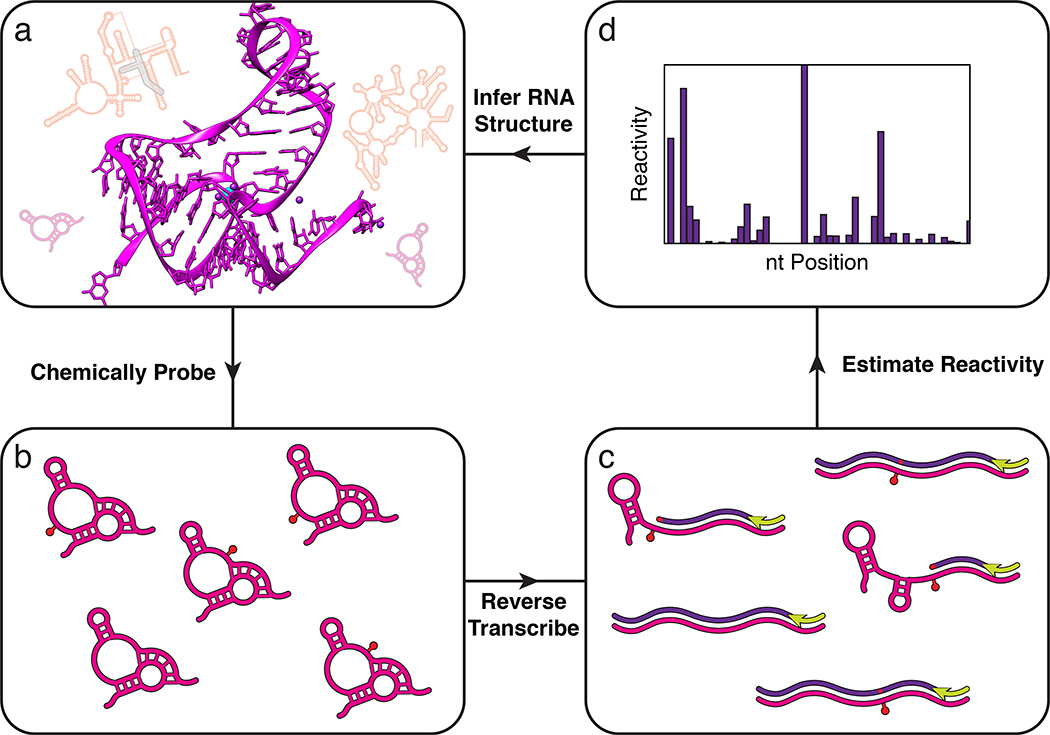
High throughput sequencing allows thousands of RNAs in complex mixtures to be structurally interrogated in a single experiment. In vitro or in vivo RNA mixtures (a) are first probed with chemical reagents that covalently modify the RNAs (red pin), or enzymes that modify the RNAs through cleavage (SI Figure 1), in a structure-dependent fashion (b) (Box 1). Modified RNAs are then converted into DNA (c) through reverse transcription that either stops or causes a mutation at each modification position (Fig. 3). The resulting pool of DNA molecules thus encode the original probe positions, and are formatted and sequenced to map the distribution of probe modifications using sequence alignment algorithms (d) (Fig. 4). The frequencies of mapped positions are used to estimate a reactivity value for each nucleotide of each RNA molecule (Box 2). High reactivities indicate nucleotides that are unstructured, while low reactivities indicate nucleotides that are constrained by RNA structures, tertiary interactions, ligand binding, or protein interactions. Reactivity information is then used to infer RNA structures (a) using a variety of computational methods.
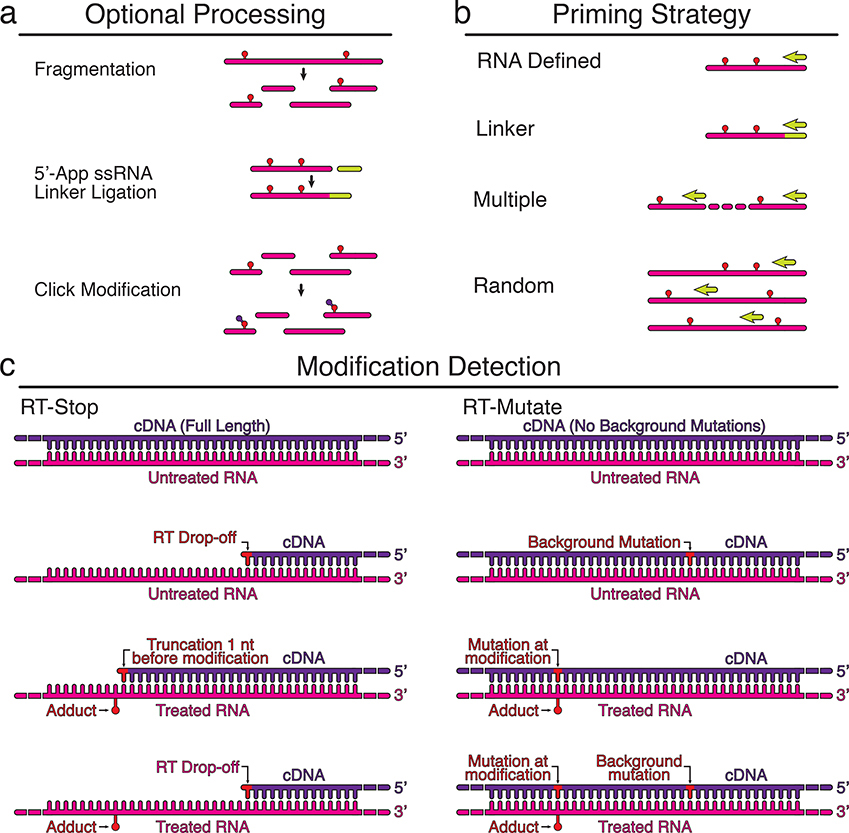
Chemical probing experiments detect the presence of RNA modifications using reverse transcription (RT). Three important variables to consider are the RNA processing steps used before RT, the RT priming strategy, and the method used to detect modifications (Table 1). Optional processing steps before RT include fragmenting the RNA, linker ligation for downstream RT priming, or modifying adducts to allow purifications (a). Several RT priming strategies can be used (b). RNA defined priming uses an RT priming site within the RNA, while linker defined primes off of the ligated linker. Multiple RT primers are used for long RNAs, and random primers can be used to tile across long RNAs, or to probe complex mixtures of RNAs and transcriptomes. Two methods are used to detect modification positions (c). In RT-stop methods, RT stops one nucleotide before the RNA modification, leading to a truncated cDNA product. In RT-mutate methods, RTs read through the modification and cause a mutation in the cDNA. Both methods result in premature truncations and background mutations in both the probed (+) and control (−) reactions that should be accounted for in downstream data analysis (Box 2).
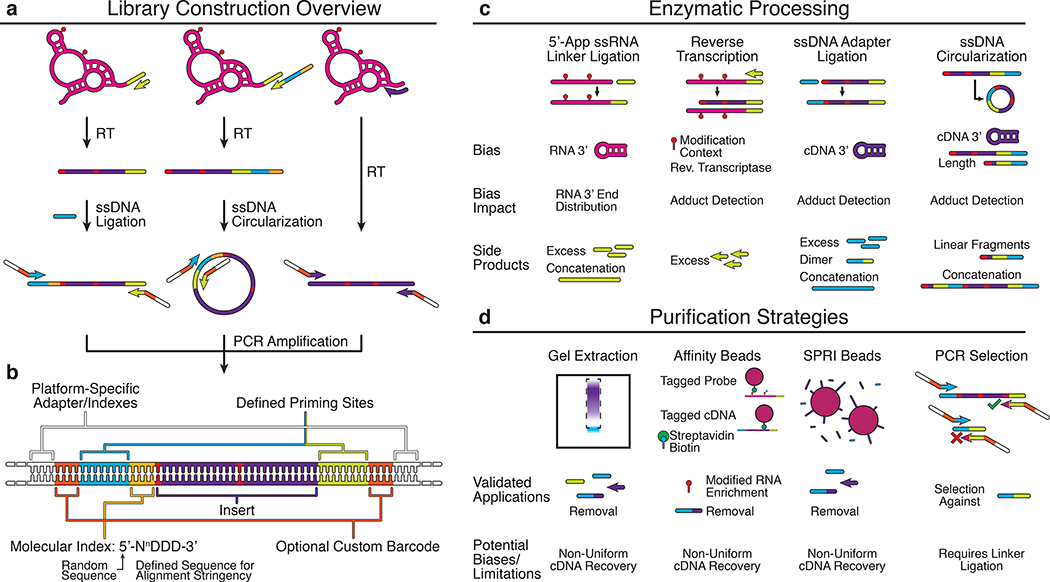
Several biochemical steps are needed in order to convert cDNA products from reverse transcription (Fig. 3) into a format compatible with sequencing instruments. After reverse transcription, a series of ligation, circularization and PCR steps are performed in different combinations to append adapter sequences needed for the sequencing process (a), and an optional set of barcodes and indices that can be used for multiplexing and downstream analysis (b). Adapter sequences are often required on both ends of the cDNA. One adapter can be conveniently included as a tail of the RT primer. The second adapter can then be ligated on through a single stranded DNA ligation, or through a circularization if the RT primer tail contained elements of both adapter sequences. Note that ligations/circularizations must be used for RT-stop techniques since each RNA modification position leads to a different cDNA end which must be mapped to recover the position. In RT-mutate methods, the presence of full length products due to RT read through allows PCR to be used to add the necessary adapter sequences. Many techniques contain additional enzymatic processing (c) and purification (d) steps throughout library preparation that get rid of excess oligonucleotides or unwanted side products that can impact downstream steps or the number of informative reads from the library. Each additional step has potential biases that must be considered when estimating reactivities (Box 2).
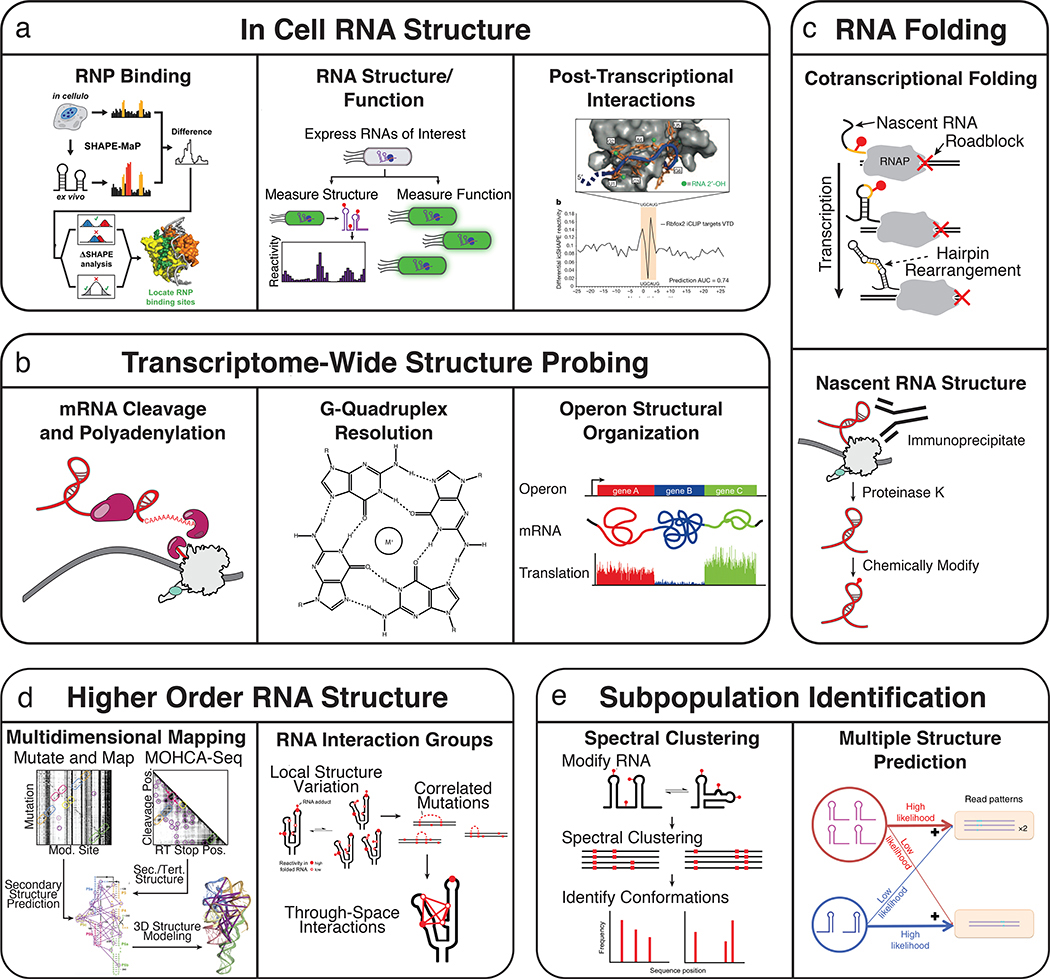
The development of high-throughput RNA structure probing methods has enabled new and diverse applications to previously inaccessible biological questions. The development of new chemical probes and analysis frameworks has enabled the characterization of in vivo RNA structure as it pertains to RNA protein interactions, RNA functional states, and post-transcriptional interactions, (a). The extension of in vivo chemical probing experiments to transcriptome wide studies has identified functional roles for RNA structure in mRNA cleavage and polyadenylation and the structural organization of operon mRNAs, and has found that G-Quadruplexes are globally unfolded in eukaryotic cells (b). The combination of high-throughput RNA structure probing and multiplexed in vitro transcription allows measurement of nascent RNA folding pathways and immunoprecipitation of Pol II has been used to isolate cellular nascent RNAs for structure probing(c). MOHCA-Seq and RING-MaP leverage the information richness of high-throughput RNA structure probing to infer higher order RNA structure (d). Structural subpopulations can be identified through spectral clustering of RING-MaP data and by multiple structure prediction using SLEQ computational analysis methods (e).
Chemical Structures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
