

Uncovering a critical period of synaptic imbalance during postnatal development of the rat visual cortex: role of brain-derived neurotrophic factor
- PMID: 30055019
- PMCID: PMC6138289
- DOI: 10.1113/JP275814
Uncovering a critical period of synaptic imbalance during postnatal development of the rat visual cortex: role of brain-derived neurotrophic factor
Abstract
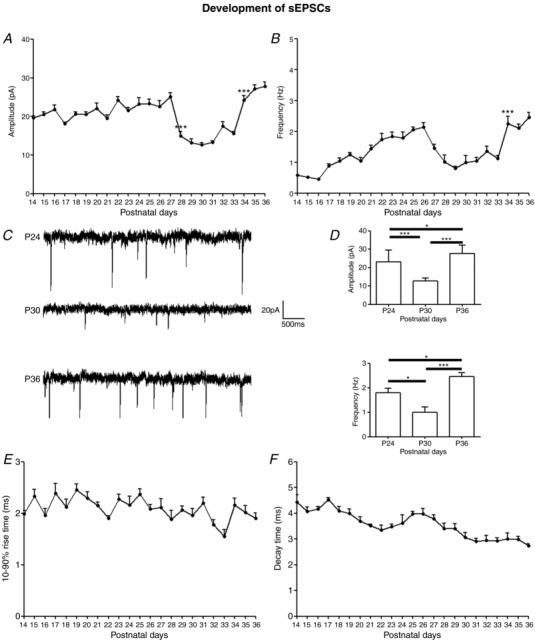
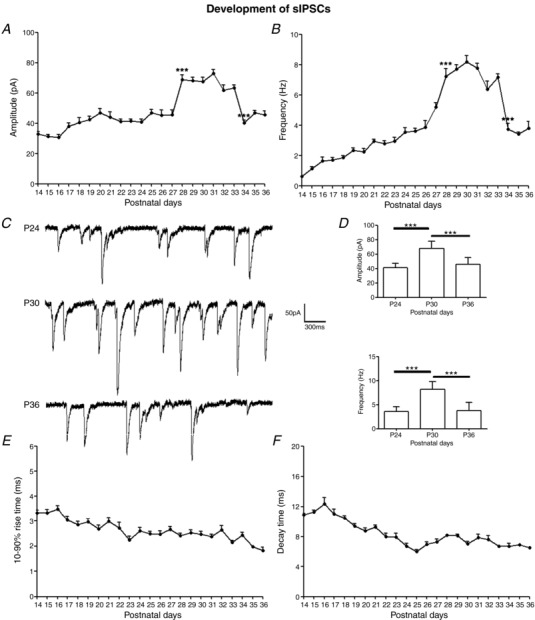
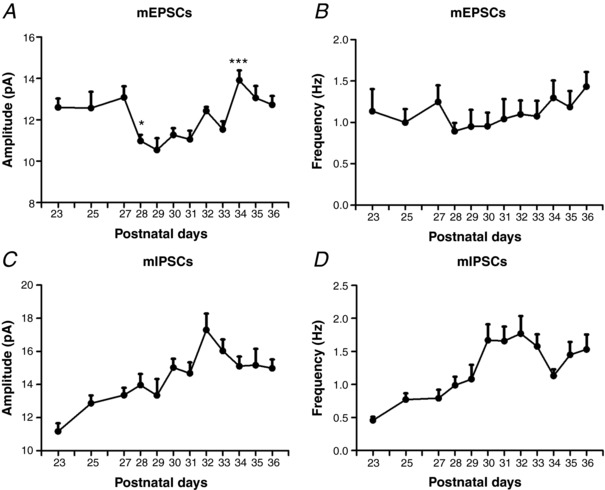
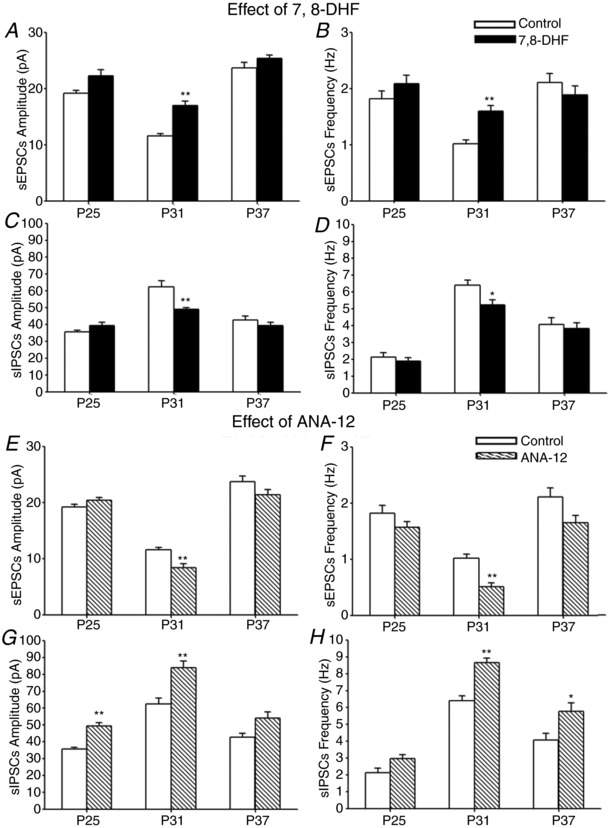
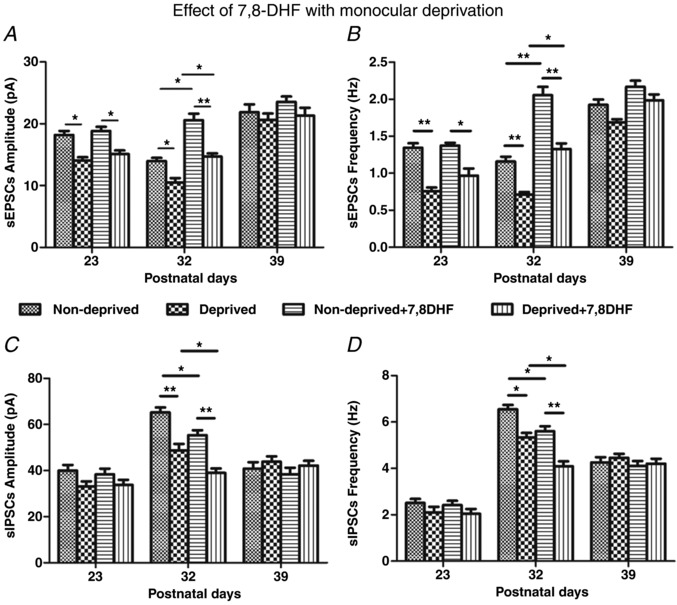
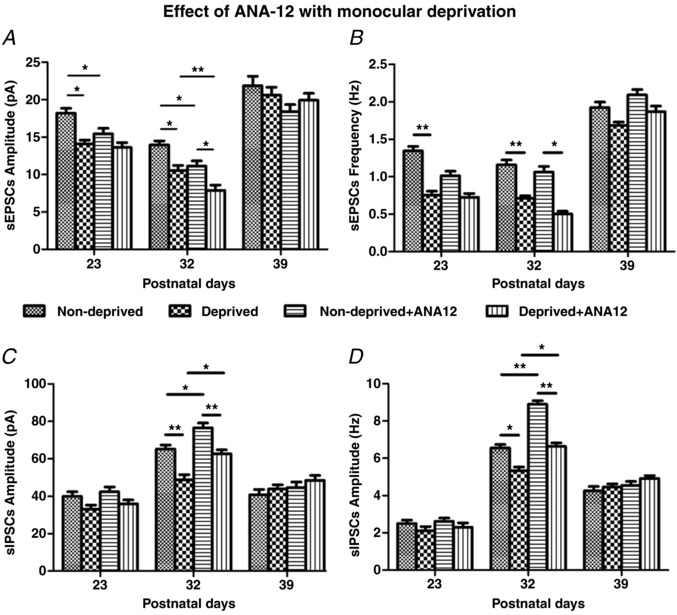
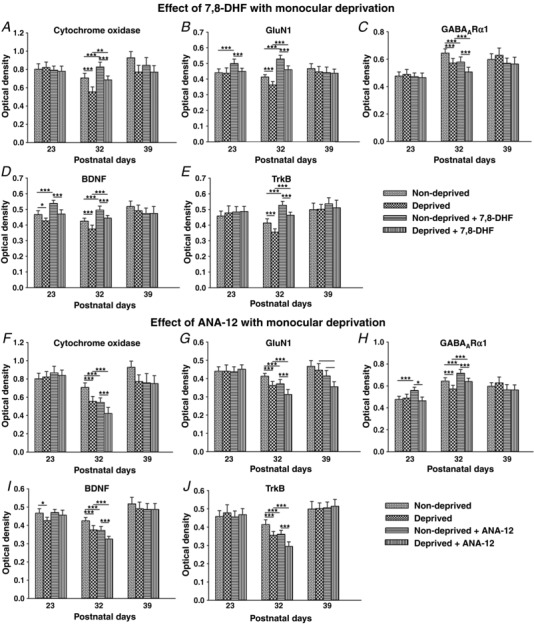
Key points: With daily electrophysiological recordings and neurochemical analysis, we uncovered a transient period of synaptic imbalance between enhanced inhibition and suppressed excitation in rat visual cortical neurons from the end of the fourth toward the end of the fifth postnatal weeks. The expression of brain-derived neurotrophic factor (BDNF), which normally enhances excitation and suppresses inhibition, was down-regulated during that time, suggesting that this may contribute to the inhibition/excitation imbalance. An agonist of the BDNF receptor tropomyosin-related kinase B (TrkB) partially reversed the imbalance, whereas a TrkB antagonist accentuated the imbalance during the transient period. Monocular lid suture during the transient period is more detrimental to the function and neurochemical properties of visual cortical neurons than before or after this period. We regard the period of synaptic imbalance as the peak critical period of vulnerability, and its existence is necessary for neurons to transition from immaturity to a more mature state of functioning.
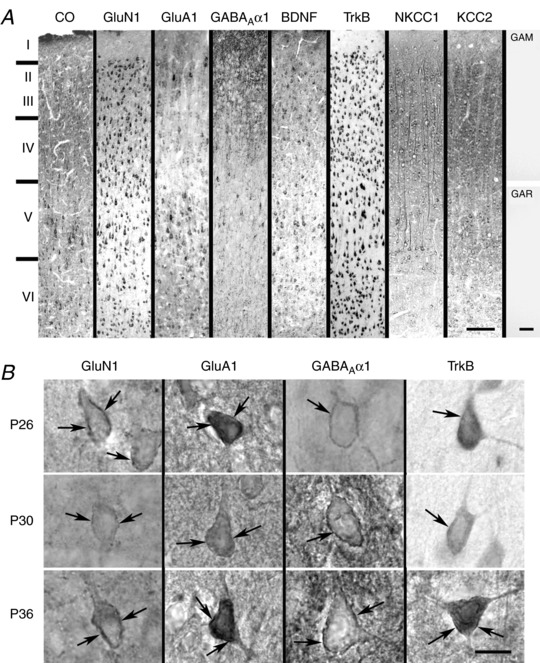
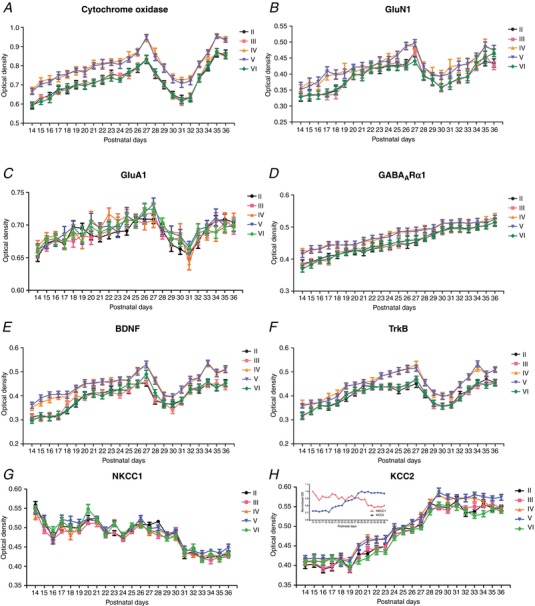
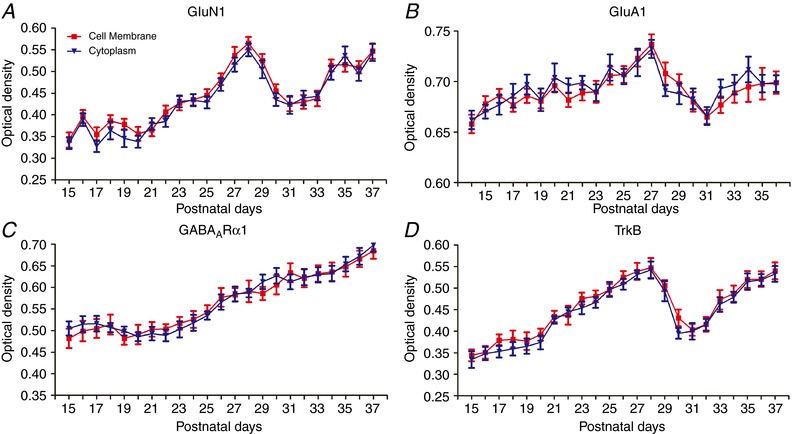
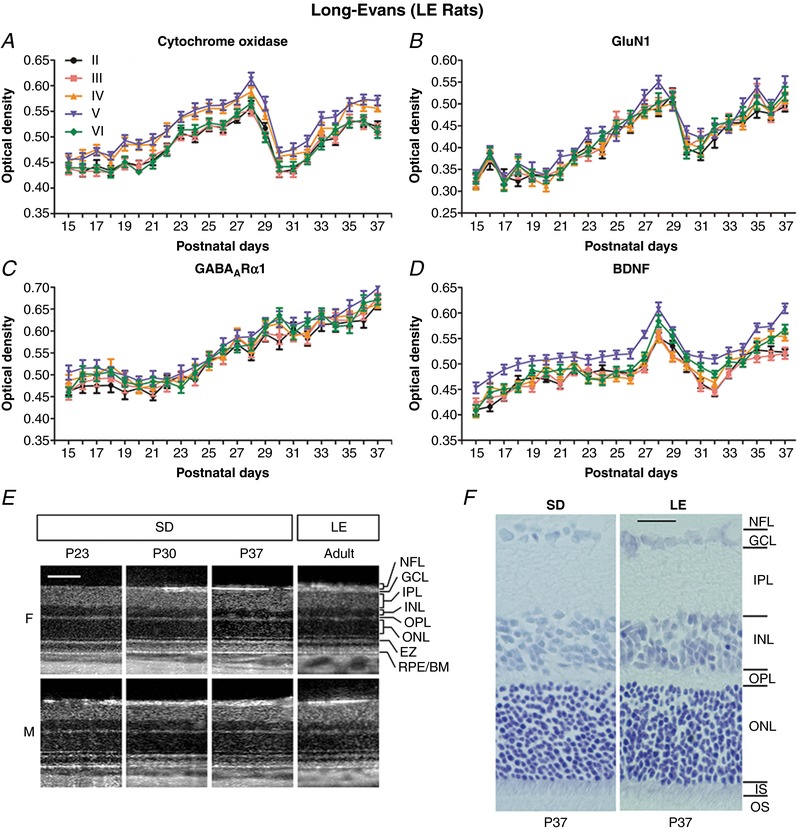
Abstract: The mammalian visual cortex is immature at birth and undergoes postnatal structural and functional adjustments. The exact timing of the vulnerable period in rodents remains unclear. The critical period is characterized by inhibitory GABAergic maturation reportedly dependent on brain-derived neurotrophic factor (BDNF). However, most of the studies were performed on experimental/transgenic animals, questioning the relationship in normal animals. The present study aimed to conduct in-depth analyses of the synaptic and neurochemical development of visual cortical neurons in normal and monocularly-deprived rats and to determine specific changes, if any, during the critical period. We found that (i) against a gradual increase in excitation and inhibition with age, a transient period of synaptic and neurochemical imbalance existed with suppressed excitation and enhanced inhibition at postnatal days 28 to 33/34; (ii) during this window, the expression of BDNF and tropomyosin-related kinase B (TrkB) receptors decreased, along with glutamatergic GluN1 and GluA1 receptors and the metabolic marker cytochrome oxidase, whereas that of GABAA Rα1 receptors continued to rise; (iii) monocular deprivation reduced both excitatory and inhibitory synaptic activity and neurochemicals mainly during this period; and (iv) in vivo TrkB agonist partially reversed the synaptic imbalance in normal and monocularly-deprived neurons during this time, whereas a TrkB antagonist accentuated the imbalance. Thus, our findings highlight a transitory period of synaptic imbalance with a negative relationship between BDNF and inhibitory GABA. This brief critical period may be necessary in transitioning from an immature to a more mature state of visual cortical functioning.
Keywords: BDNF; critical period; immunohistochemistry; monocular deprivation; patch-clamp recording; visual cortex.
© 2018 The Authors. The Journal of Physiology © 2018 The Physiological Society.
Figures

Comment in
-
Transient downregulation of BDNF is required for GABAergic maturation in rat primary visual cortex.J Physiol. 2019 Feb;597(3):673-675. doi: 10.1113/JP277386. Epub 2018 Dec 11. J Physiol. 2019. PMID: 30506569 Free PMC article. No abstract available.
References
-
- Alvarez‐Leefmans FJ, León‐Olea M, Mendoza‐Sotelo J, Alvarez FJ, Antón B & Garduño R (2001). Immunolocalization of the Na(+)‐K(+)‐2Cl(–) cotransporter in peripheral nervous tissue of vertebrates. Neurosci 104, 569–582. - PubMed
-
- Ben‐Ari Y (2002). Excitatory actions of GABA during development: the nature of the nurture. Nat Rev Neurosci 3, 728–739. - PubMed
-
- Berger A, Cavallero S, Dominguez E, Barber P, Simonutti M, Sahel JA, Sennlaub F, Raoul W, Paques M & Bemelmans AP (2014). Spectral‐domain optical coherence tomography of the rodent eye: highlighting layers of the outer retina using signal averaging and comparison with histology. PLoS ONE 9, e96494. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources