

Progenitor Hyperpolarization Regulates the Sequential Generation of Neuronal Subtypes in the Developing Neocortex
- PMID: 30057116
- PMCID: PMC6545245
- DOI: 10.1016/j.cell.2018.06.036
Progenitor Hyperpolarization Regulates the Sequential Generation of Neuronal Subtypes in the Developing Neocortex
Abstract
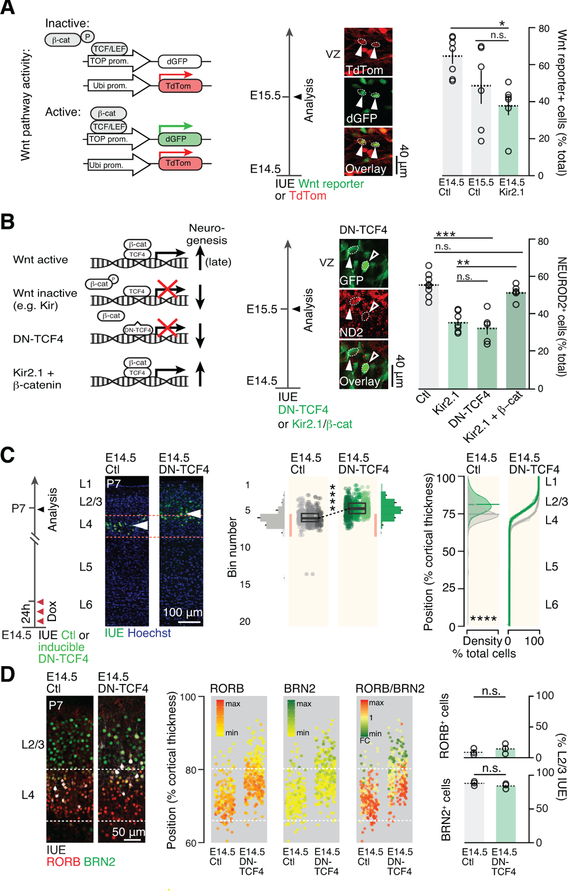
During corticogenesis, ventricular zone progenitors sequentially generate distinct subtypes of neurons, accounting for the diversity of neocortical cells and the circuits they form. While activity-dependent processes are critical for the differentiation and circuit assembly of postmitotic neurons, how bioelectrical processes affect nonexcitable cells, such as progenitors, remains largely unknown. Here, we reveal that, in the developing mouse neocortex, ventricular zone progenitors become more hyperpolarized as they generate successive subtypes of neurons. Experimental in vivo hyperpolarization shifted the transcriptional programs and division modes of these progenitors to a later developmental status, with precocious generation of intermediate progenitors and a forward shift in the laminar, molecular, morphological, and circuit features of their neuronal progeny. These effects occurred through inhibition of the Wnt-beta-catenin signaling pathway by hyperpolarization. Thus, during corticogenesis, bioelectric membrane properties are permissive for specific molecular pathways to coordinate the temporal progression of progenitor developmental programs and thus neocortical neuron diversity.
Keywords: cortical development; membrane potential; neuronal diversity; progenitors.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
No competing interests to declare.
Figures


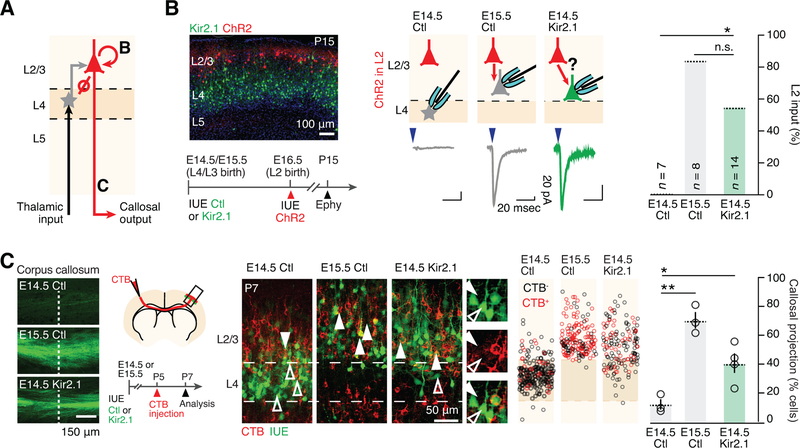
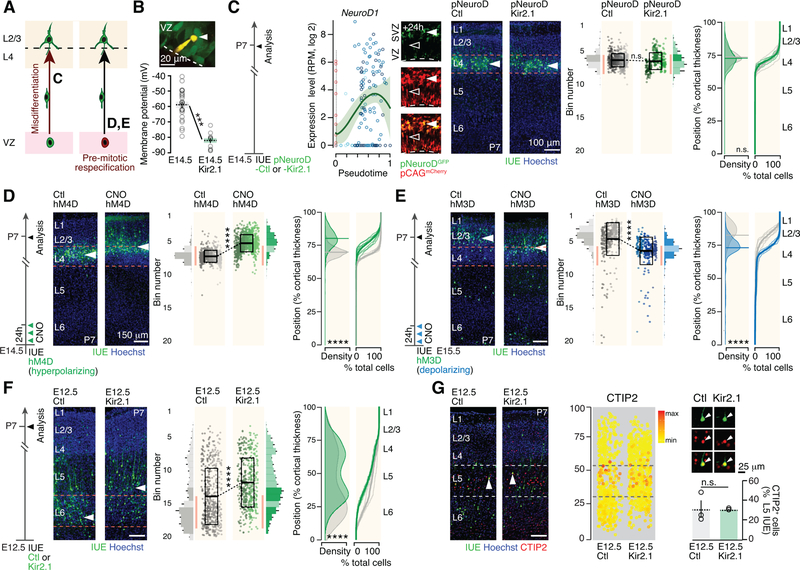
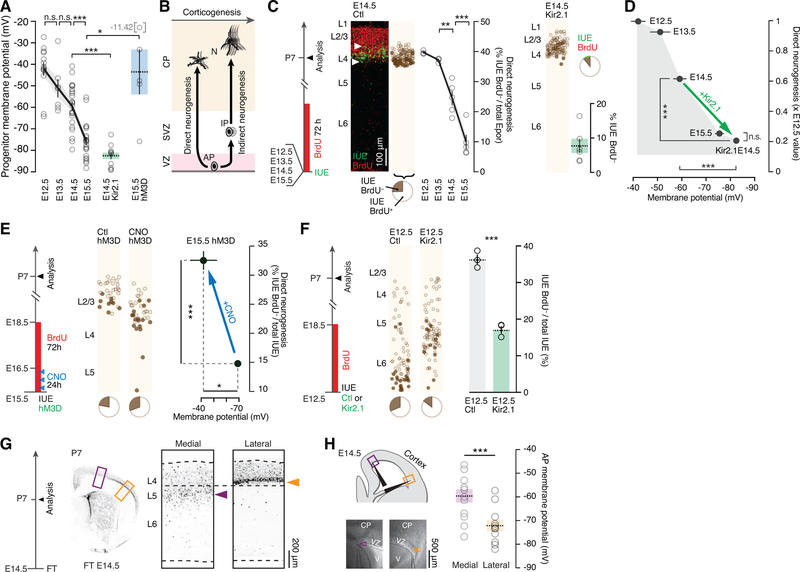
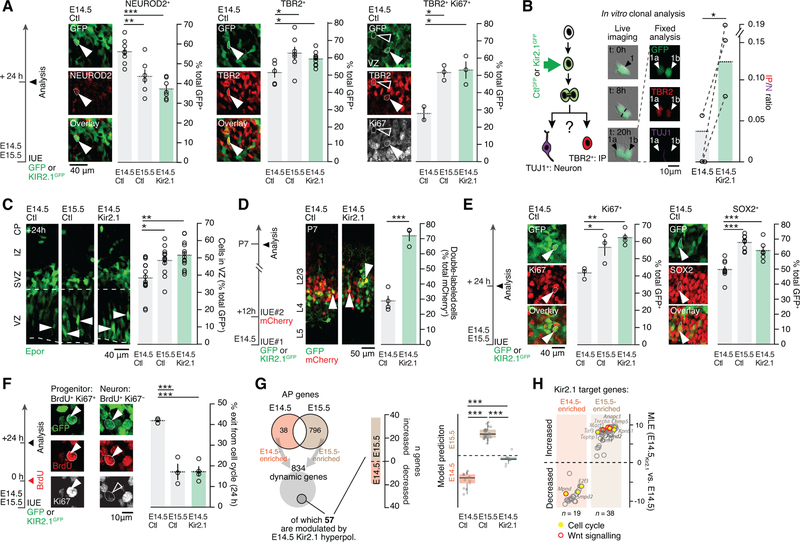
Comment in
-
Hands of (cell) fate.Nat Rev Neurosci. 2018 Sep;19(9):518. doi: 10.1038/s41583-018-0050-z. Nat Rev Neurosci. 2018. PMID: 30089889 No abstract available.
-
An Electric Take on Neural Fate and Cortical Development.Dev Cell. 2019 Jan 7;48(1):1-2. doi: 10.1016/j.devcel.2018.12.014. Dev Cell. 2019. PMID: 30620896
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
