

Small RNAs Gained during Epididymal Transit of Sperm Are Essential for Embryonic Development in Mice
- PMID: 30057276
- PMCID: PMC6103825
- DOI: 10.1016/j.devcel.2018.06.024
Small RNAs Gained during Epididymal Transit of Sperm Are Essential for Embryonic Development in Mice
Abstract
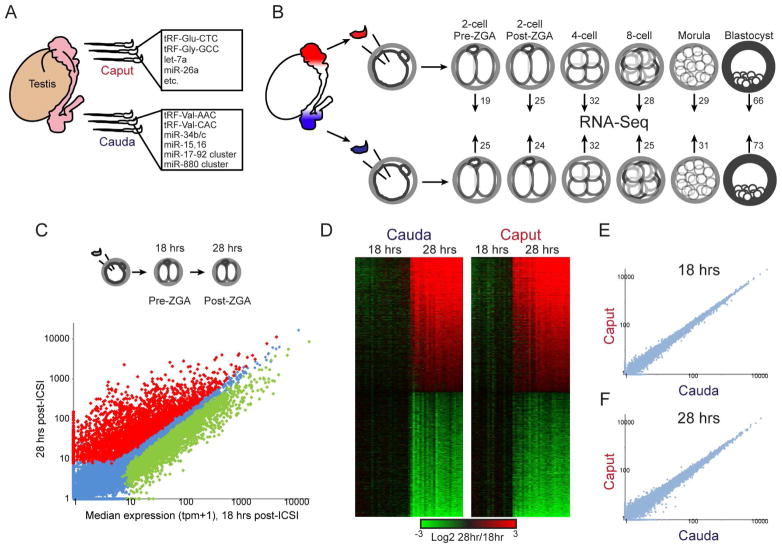
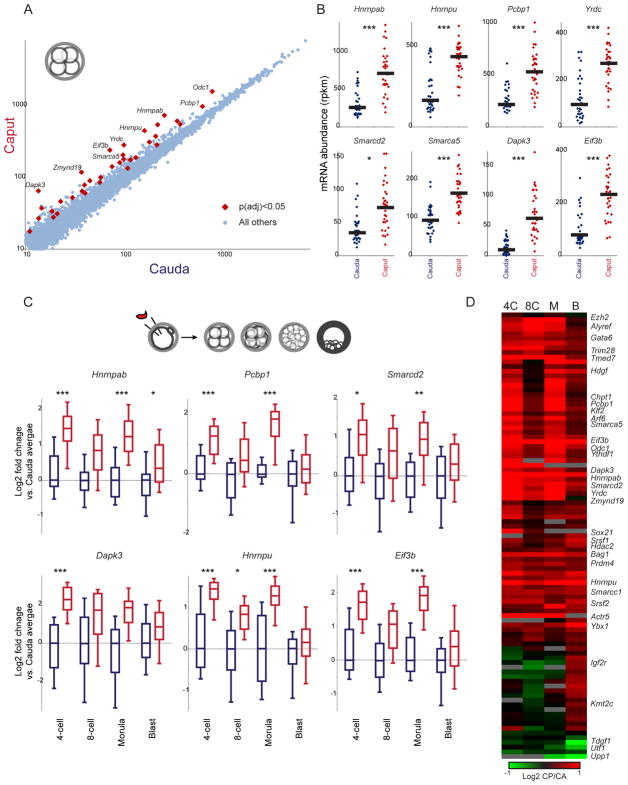
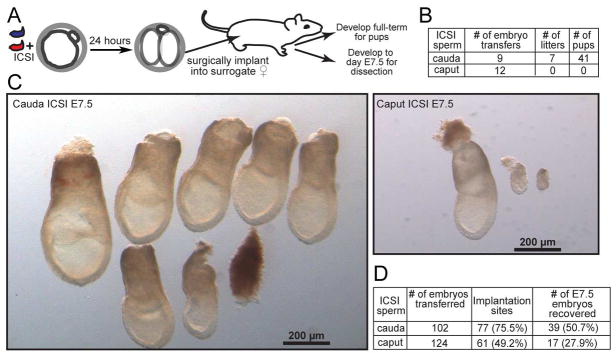
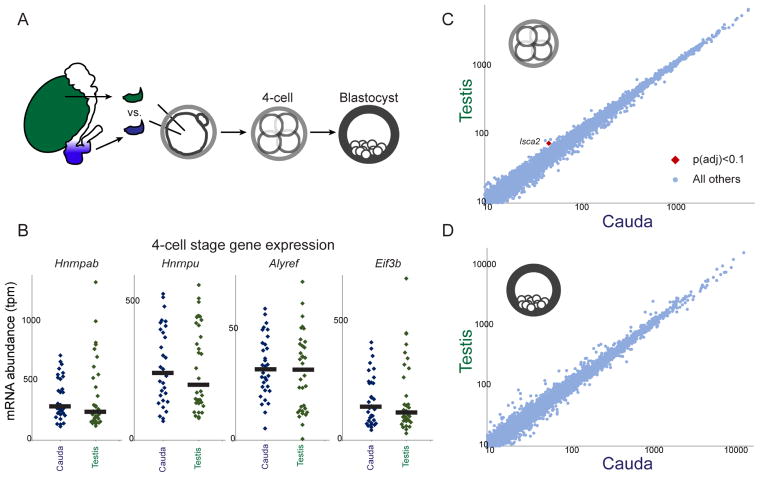
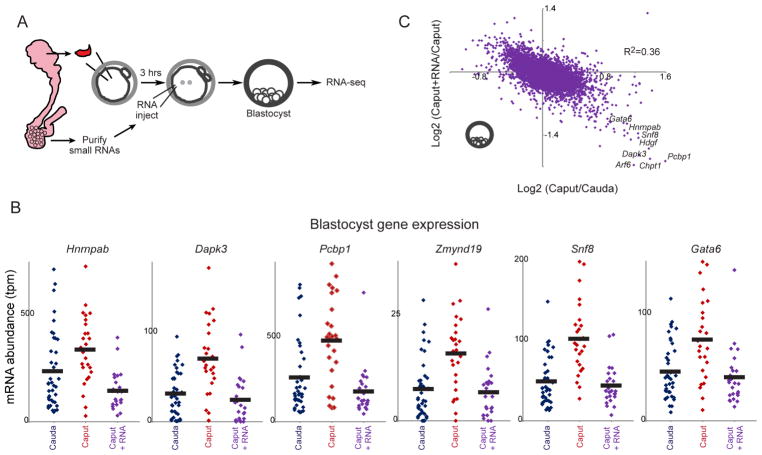
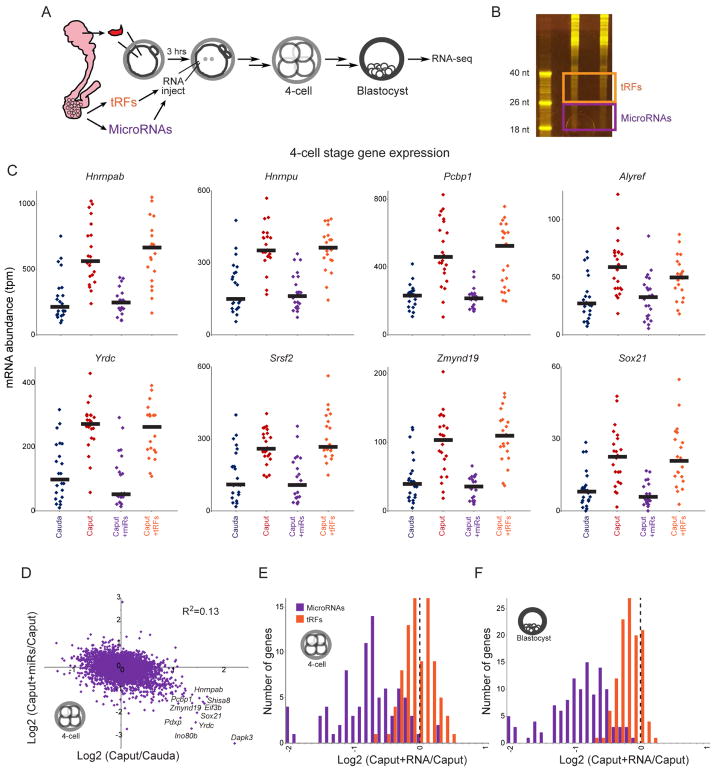
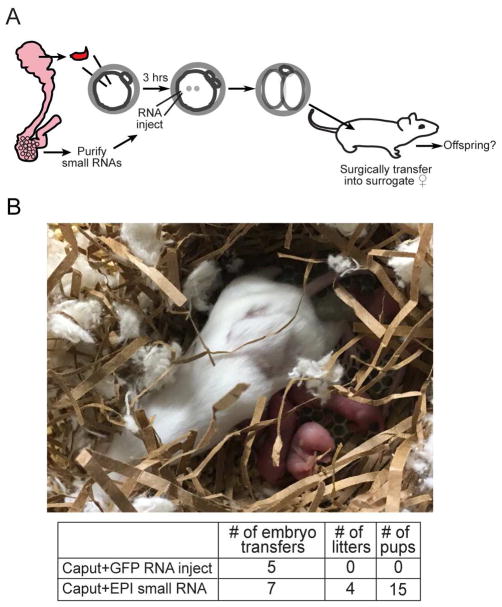
The small RNA payload of mammalian sperm undergoes dramatic remodeling during development, as several waves of microRNAs and tRNA fragments are shipped to sperm during post-testicular maturation in the epididymis. Here, we take advantage of this developmental process to probe the function of the sperm RNA payload in preimplantation development. We generated zygotes via intracytoplasmic sperm injection (ICSI) using sperm obtained from the proximal (caput) versus distal (cauda) epididymis and then characterized the development of the resulting embryos. Embryos generated using caput sperm significantly overexpress multiple regulatory factors throughout preimplantation development, subsequently implant inefficiently, and fail soon after implantation. Remarkably, microinjection of purified cauda-specific small RNAs into caput-derived embryos not only completely rescued preimplantation molecular defects but also suppressed the post-implantation embryonic lethality phenotype. These findings reveal an essential role for small RNA remodeling during post-testicular maturation of mammalian sperm and identify a specific preimplantation gene expression program responsive to sperm-delivered microRNAs.
Keywords: epigenetics; preimplantation development; small RNAs; spermatogenesis.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Dynamic RNAs in sperm shape embryo development.Nat Rev Genet. 2018 Oct;19(10):591. doi: 10.1038/s41576-018-0042-x. Nat Rev Genet. 2018. PMID: 30069060 No abstract available.
-
Caput Epididymidal Mouse Sperm Support Full Development.Dev Cell. 2019 Jul 1;50(1):5-6. doi: 10.1016/j.devcel.2019.05.012. Dev Cell. 2019. PMID: 31265812 No abstract available.
-
MicroRNAs Absent in Caput Sperm Are Required for Normal Embryonic Development.Dev Cell. 2019 Jul 1;50(1):7-8. doi: 10.1016/j.devcel.2019.06.007. Dev Cell. 2019. PMID: 31265813 No abstract available.
-
Both Cauda and Caput Epididymal Sperm Are Capable of Supporting Full-Term Development in FVB and CD-1 Mice.Dev Cell. 2020 Dec 21;55(6):675-676. doi: 10.1016/j.devcel.2020.11.022. Dev Cell. 2020. PMID: 33352141 Free PMC article. No abstract available.
-
Sperm Head Preparation and Genetic Background Affect Caput Sperm ICSI Embryo Viability: Cauda-Enriched miRNAs Only Essential in Specific Conditions.Dev Cell. 2020 Dec 21;55(6):677-678. doi: 10.1016/j.devcel.2020.11.021. Dev Cell. 2020. PMID: 33352142 No abstract available.
References
-
- Belleannee C, Calvo E, Caballero J, Sullivan R. Epididymosomes convey different repertoires of microRNAs throughout the bovine epididymis. Biology of reproduction. 2013;89:30. - PubMed
-
- Chen Q, Yan M, Cao Z, Li X, Zhang Y, Shi J, Feng GH, Peng H, Zhang X, Zhang Y, et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science (New York, N Y. 2016;351:397–400. - PubMed
-
- Donkin I, Versteyhe S, Ingerslev LR, Qian K, Mechta M, Nordkap L, Mortensen B, Appel EV, Jorgensen N, Kristiansen VB, et al. Obesity and Bariatric Surgery Drive Epigenetic Variation of Spermatozoa in Humans. Cell metabolism. 2016;23:369–378. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
