

An APOBEC3A-Cas9 base editor with minimized bystander and off-target activities
- PMID: 30059493
- PMCID: PMC6181770
- DOI: 10.1038/nbt.4199
An APOBEC3A-Cas9 base editor with minimized bystander and off-target activities
Abstract
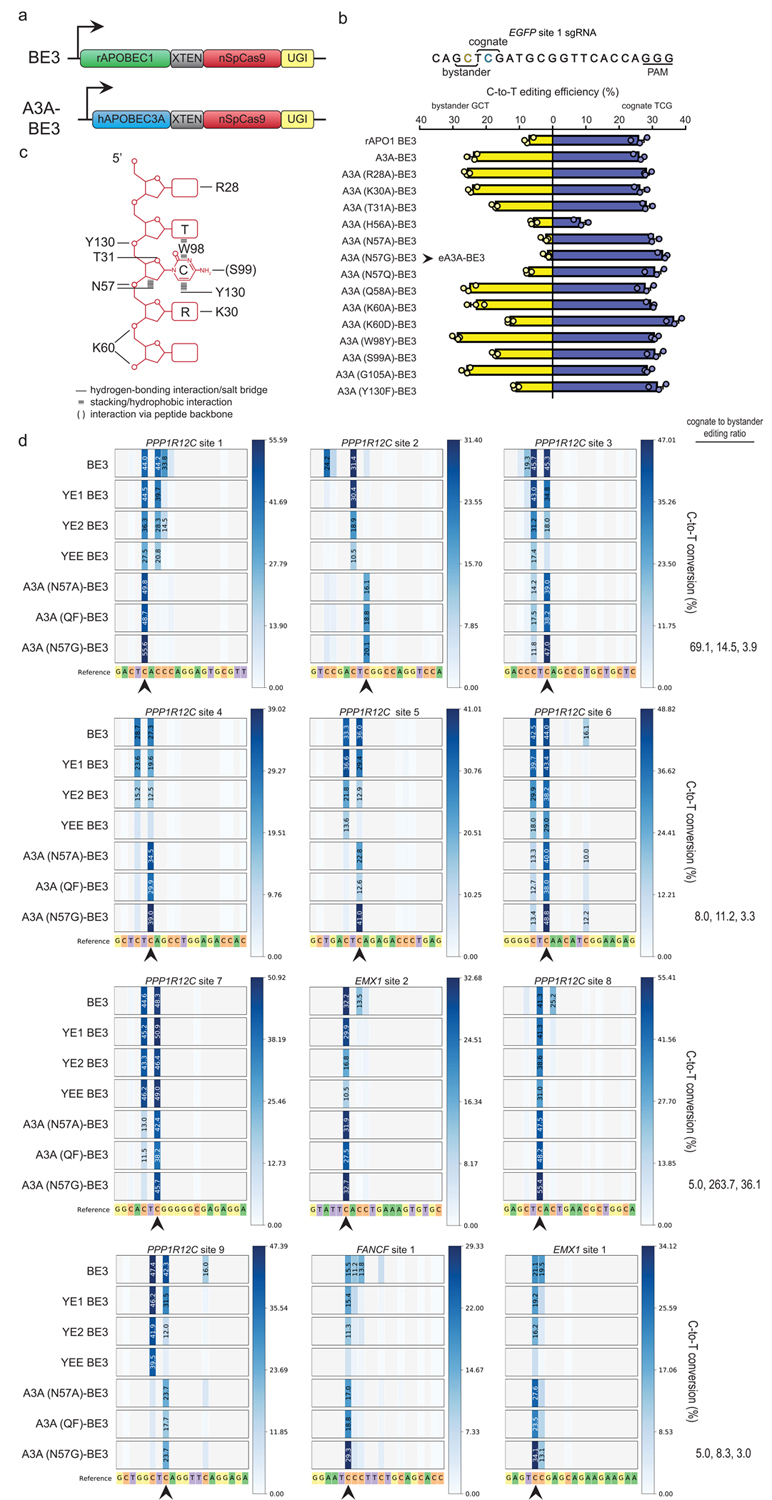
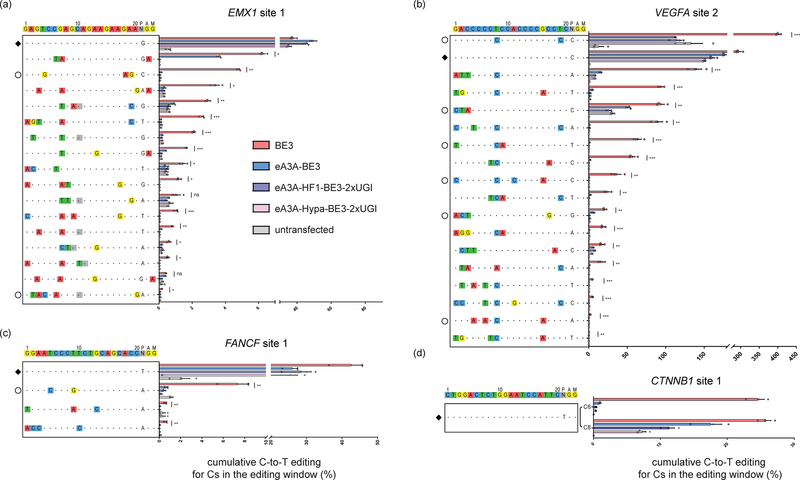
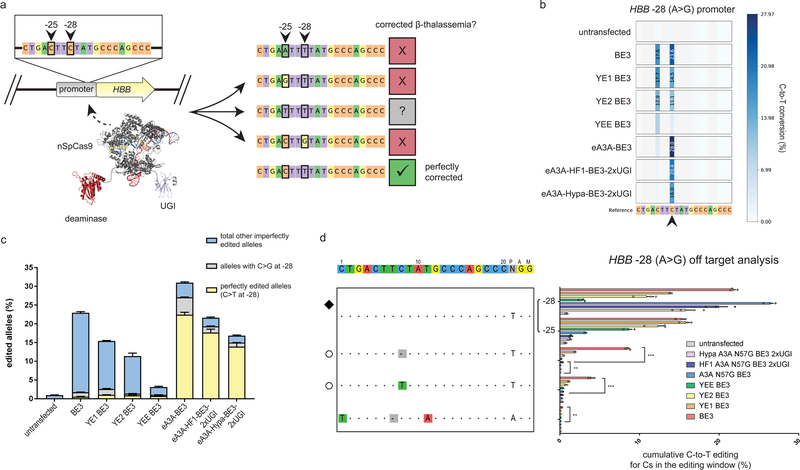
Base editor technology, which uses CRISPR-Cas9 to direct cytidine deaminase enzymatic activity to specific genomic loci, enables the highly efficient introduction of precise cytidine-to-thymidine DNA alterations. However, existing base editors create unwanted C-to-T alterations when more than one C is present in the enzyme's five-base-pair editing window. Here we describe a strategy for reducing bystander mutations using an engineered human APOBEC3A (eA3A) domain, which preferentially deaminates cytidines in specific motifs according to a TCR>TCY>VCN hierarchy. In direct comparisons with the widely used base editor 3 (BE3) fusion in human cells, our eA3A-BE3 fusion exhibits similar activities on cytidines in TC motifs but greatly reduced editing on cytidines in other sequence contexts. eA3A-BE3 corrects a human β-thalassemia promoter mutation with much higher (>40-fold) precision than BE3. We also demonstrate that eA3A-BE3 shows reduced mutation frequencies on known off-target sites of BE3, even when targeting promiscuous homopolymeric sites.
Conflict of interest statement
J.M.G. is currently a full-time employee of and holds equity in Beam Therapeutics. J.K.J. has financial interests in Beam Therapeutics, Editas Medicine, Monitor Biotechnologies, Pairwise Plants, Poseida Therapeutics, and Transposagen Biopharmaceuticals. J.K.J.’s interests were reviewed and are managed by Massachusetts General Hospital and Partners HealthCare in accordance with their conflict of interest policies. J.M.G. and J.K.J. are inventors on a patent application that has been filed for engineered sequence-specific deaminase domains in base editor architectures.
Figures
Comment in
-
Better base editors.Nat Methods. 2018 Oct;15(10):763. doi: 10.1038/s41592-018-0154-4. Nat Methods. 2018. PMID: 30275576 No abstract available.
References
-
- Nishida K et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science 353, (2016). - PubMed
-
- Kim K et al. Highly efficient RNA-guided base editing in mouse embryos. Nat. Biotechnol 35, 435–437 (2017). - PubMed
-
- Shimatani Z et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol 35, 441–443 (2017). - PubMed
METHODS-ONLY REFERENCES
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
