

Sex-Dimorphic and Sex Hormone-Dependent Role of Steroid Sulfatase in Adipose Inflammation and Energy Homeostasis
- PMID: 30060148
- PMCID: PMC6112598
- DOI: 10.1210/en.2018-00531
Sex-Dimorphic and Sex Hormone-Dependent Role of Steroid Sulfatase in Adipose Inflammation and Energy Homeostasis
Abstract
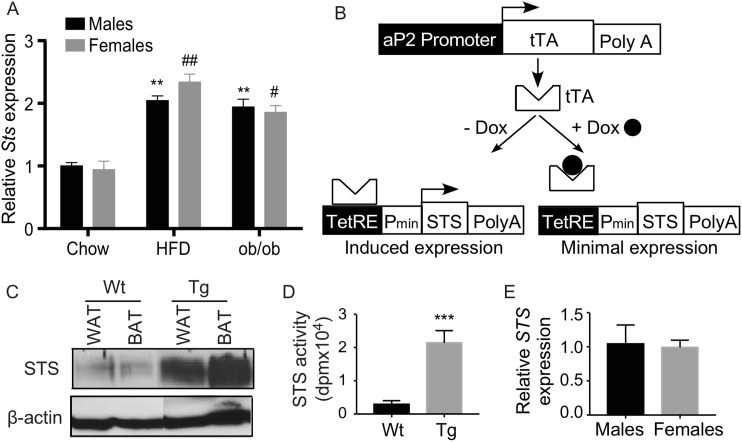
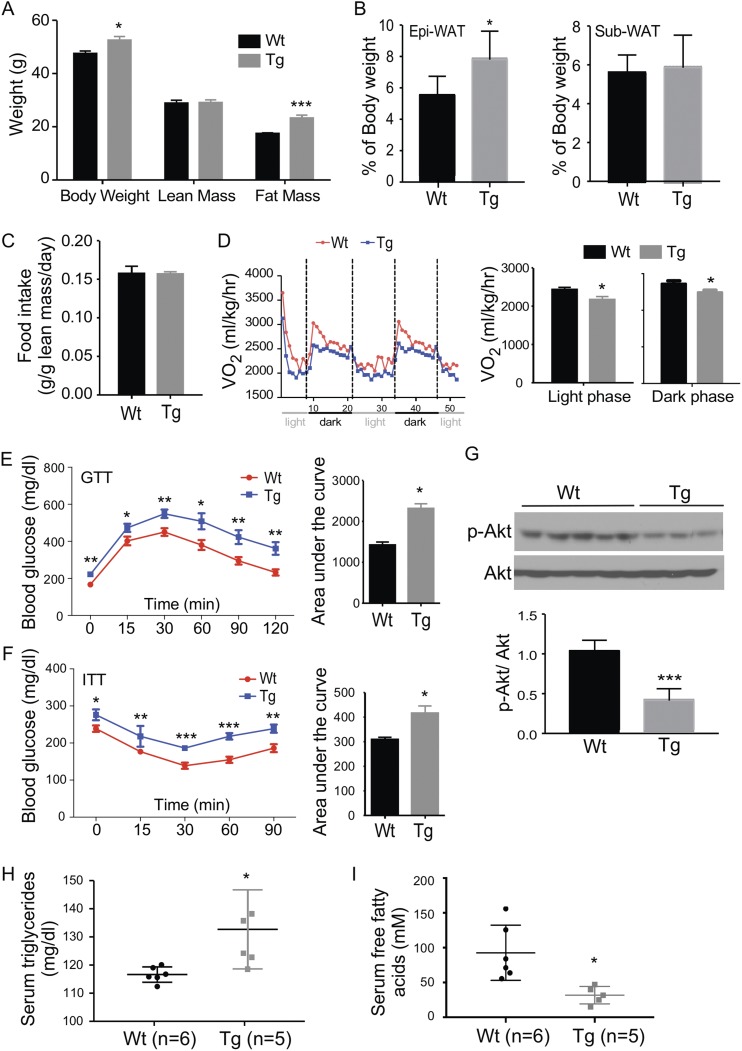
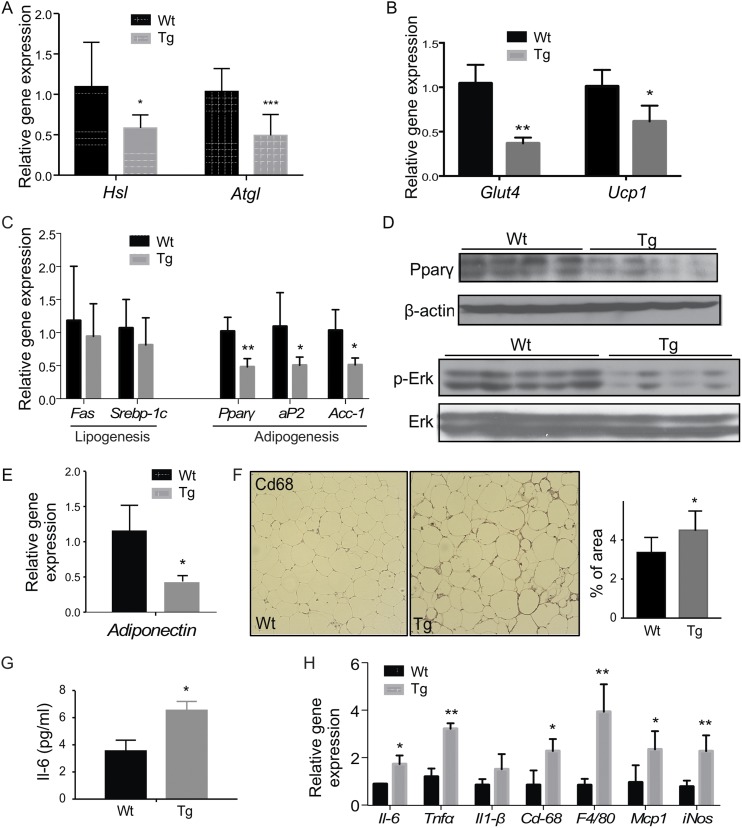
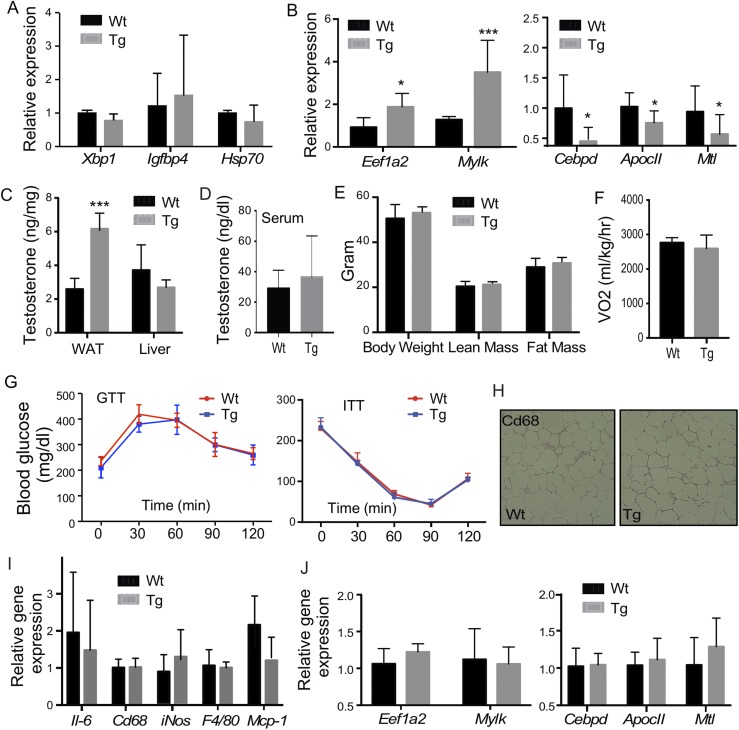
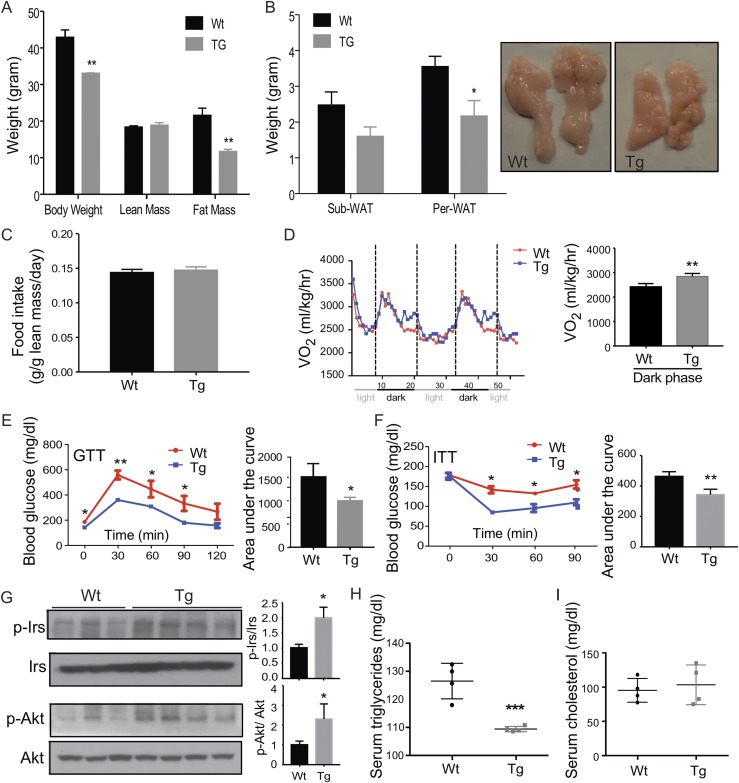
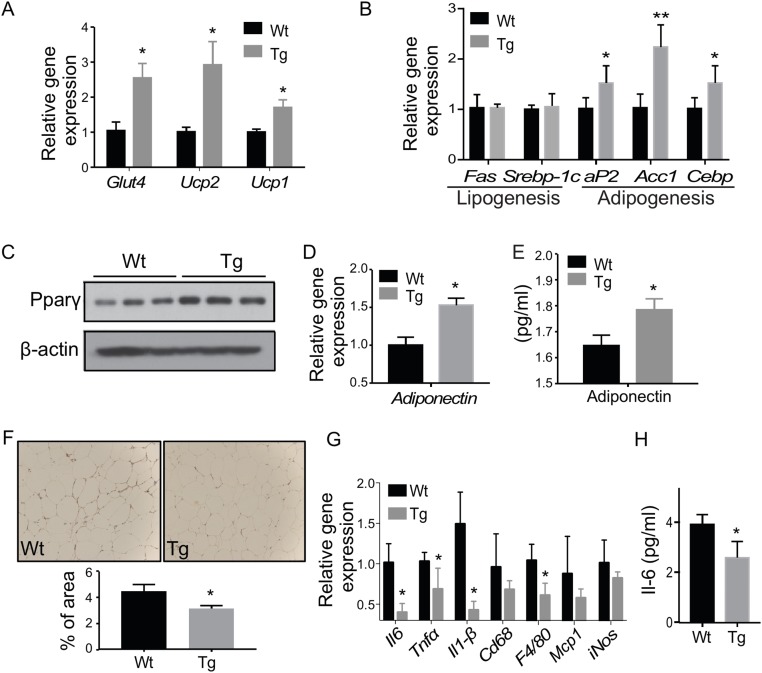
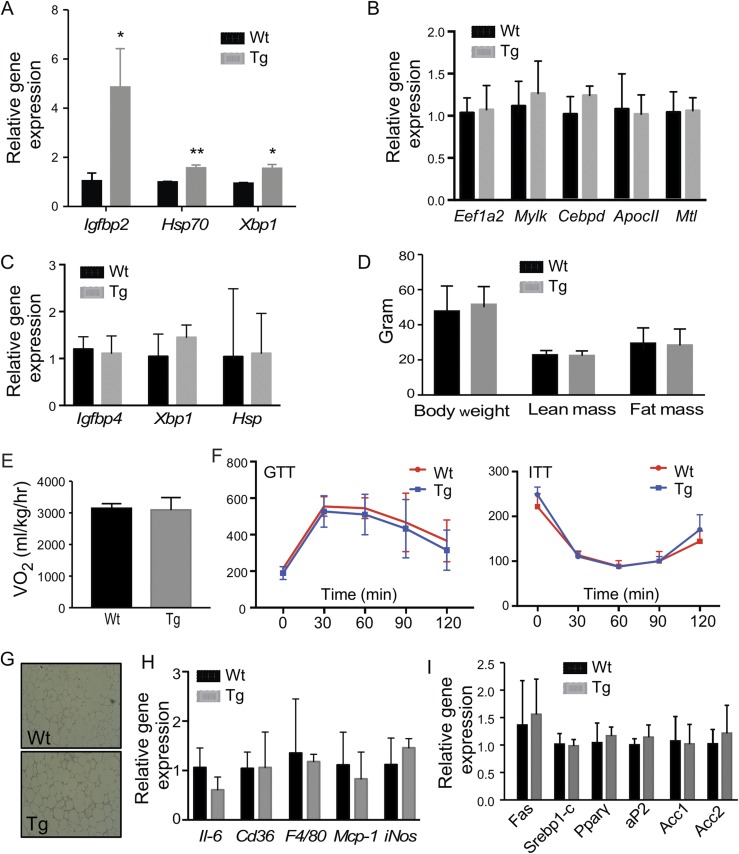
Steroid sulfatase (STS), a desulfating enzyme that converts steroid sulfates to hormonally active steroids, plays an important role in the homeostasis of sex hormones. STS is expressed in the adipose tissue of both male and female mice, but the role of STS in the development and function of adipose tissue remains largely unknown. In this report, we show that the adipose expression of Sts was induced in the high-fat diet (HFD) and ob/ob models of obesity and type 2 diabetes. Transgenic overexpression of the human STS in the adipose tissue of male mice exacerbated the HFD-induced metabolic phenotypes, including increased body weight gain and fat mass, and worsened insulin sensitivity, glucose tolerance, and energy expenditure, which were accounted for by adipocyte hypertrophy, increased adipose inflammation, and dysregulation of adipogenesis. The metabolic harm of the STS transgene appeared to have resulted from increased androgen activity in the adipose tissue, and castration abolished most of the phenotypes. Interestingly, the transgenic effects were sex specific, because the HFD-fed female STS transgenic mice exhibited improved metabolic functions, which were associated with attenuated adipose inflammation. The metabolic benefit of the STS transgene in female mice was accounted for by increased estrogenic activity in the adipose tissue, whereas such benefit was abolished upon ovariectomy. Our results revealed an essential role of the adipose STS in energy homeostasis in sex- and sex hormone-dependent manner. The adipose STS may represent a therapeutic target for the management of obesity and type 2 diabetes.
Figures
References
-
- Oh JY, Barrett-Connor E, Wedick NM, Wingard DL; Rancho Bernardo Study . Endogenous sex hormones and the development of type 2 diabetes in older men and women: the Rancho Bernardo study. Diabetes Care. 2002;25(1):55–60. - PubMed
-
- Carr MC. The emergence of the metabolic syndrome with menopause. J Clin Endocrinol Metab. 2003;88(6):2404–2411. - PubMed
-
- Smith EP, Boyd J, Frank GR, Takahashi H, Cohen RM, Specker B, Williams TC, Lubahn DB, Korach KS. Estrogen resistance caused by a mutation in the estrogen-receptor gene in a man. N Engl J Med. 1994;331(16):1056–1061. - PubMed
-
- Morishima A, Grumbach MM, Simpson ER, Fisher C, Qin K. Aromatase deficiency in male and female siblings caused by a novel mutation and the physiological role of estrogens. J Clin Endocrinol Metab. 1995;80(12):3689–3698. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
