

Protein motion in the nucleus: from anomalous diffusion to weak interactions
- PMID: 30065106
- PMCID: PMC6103463
- DOI: 10.1042/BST20170310
Protein motion in the nucleus: from anomalous diffusion to weak interactions
Abstract
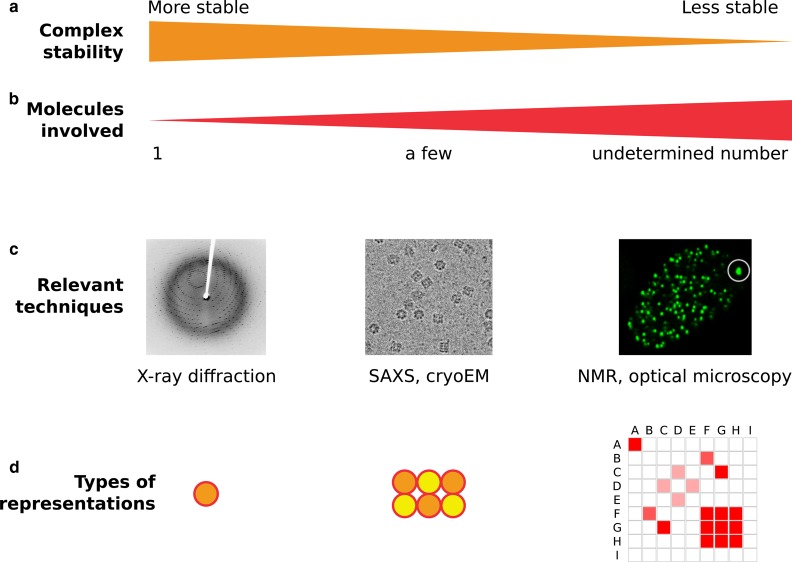
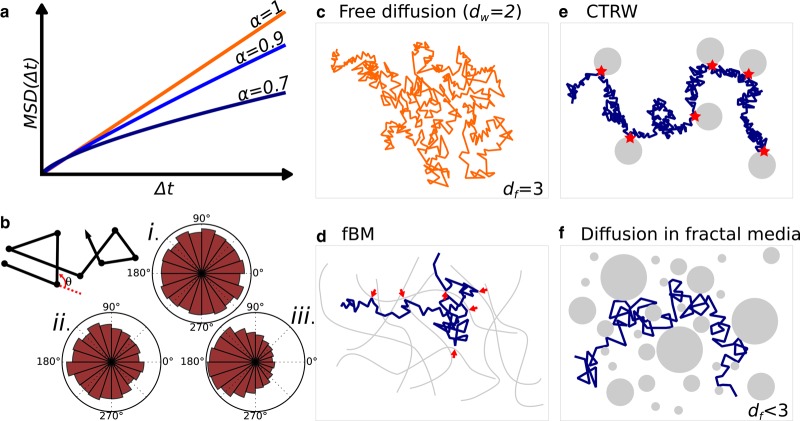
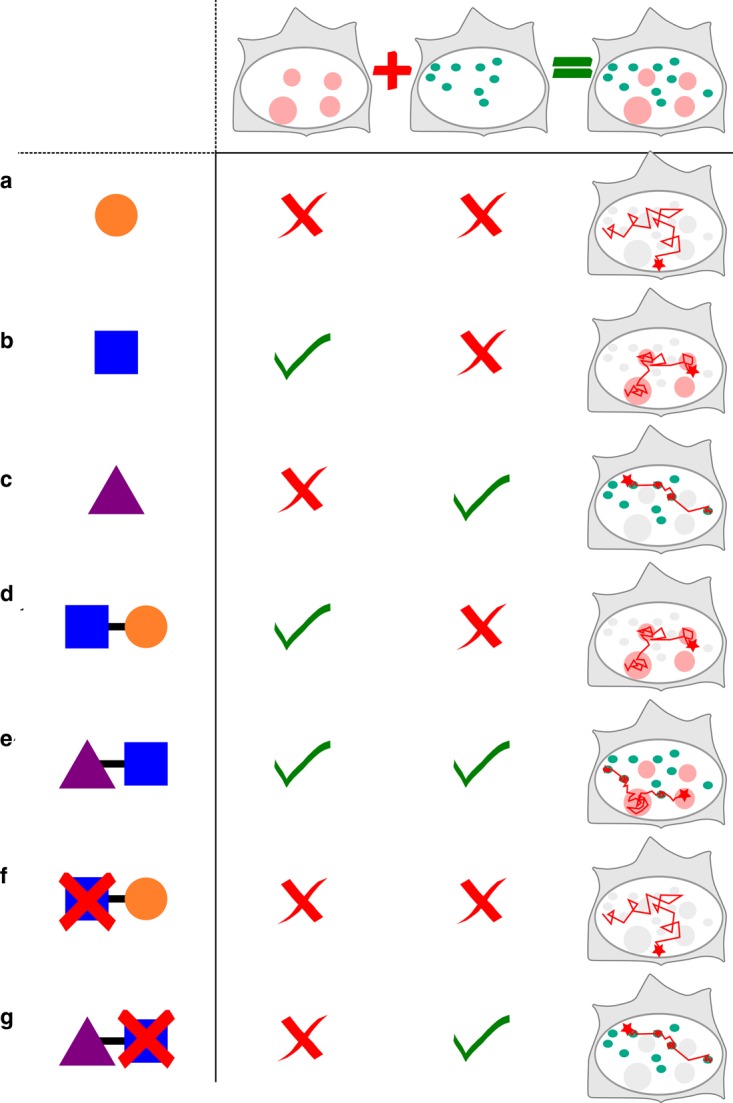
Understanding how transcription factors (TFs) regulate mammalian gene expression in space and time is a central topic in biology. To activate a gene, a TF has first to diffuse in the available space of the nucleus until it reaches a target DNA sequence or protein (target site). This eventually results in the recruitment of the whole transcriptional machinery. All these processes take place in the mammalian nucleoplasm, a highly organized and dynamic environment, in which some complexes transiently assemble and break apart, whereas others appear more stable. This diversity of dynamic behaviors arises from the number of biomolecules that make up the nucleoplasm and their pairwise interactions. Indeed, interactions energies that span several orders of magnitude, from covalent bounds to transient and dynamic interactions, can shape nuclear landscapes. Thus, the nuclear environment determines how frequently and how fast a TF contacts its target site, and it indirectly regulates gene expression. How exactly transient interactions are involved in the regulation of TF diffusion is unclear, but are reflected by live cell imaging techniques, including single-particle tracking (SPT). Overall, the macroscopic result of these microscopic interactions is almost always anomalous diffusion, a phenomenon widely studied and modeled. Here, we review the connections between the anomalous diffusion of a TF observed by SPT and the microscopic organization of the nucleus, including recently described topologically associated domains and dynamic phase-separated compartments. We propose that anomalous diffusion found in SPT data result from weak and transient interactions with dynamic nuclear substructures, and that SPT data analysis would benefit from a better description of such structures.
Keywords: molecular interactions; nuclear transport; protein motion; transcription factor.
© 2018 The Author(s).
Conflict of interest statement
The Author declare that there are no competing interests associated with this manuscript.
Figures
References
-
- von Smoluchowski M. (1906) Zur kinetischen theorie der brownschen molekularbewegung und der suspensionen. Ann. Phys. 326, 756–780 10.1002/andp.19063261405 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous