

COMT1 Silencing Aggravates Heat Stress-Induced Reduction in Photosynthesis by Decreasing Chlorophyll Content, Photosystem II Activity, and Electron Transport Efficiency in Tomato
- PMID: 30065736
- PMCID: PMC6056654
- DOI: 10.3389/fpls.2018.00998
COMT1 Silencing Aggravates Heat Stress-Induced Reduction in Photosynthesis by Decreasing Chlorophyll Content, Photosystem II Activity, and Electron Transport Efficiency in Tomato
Abstract
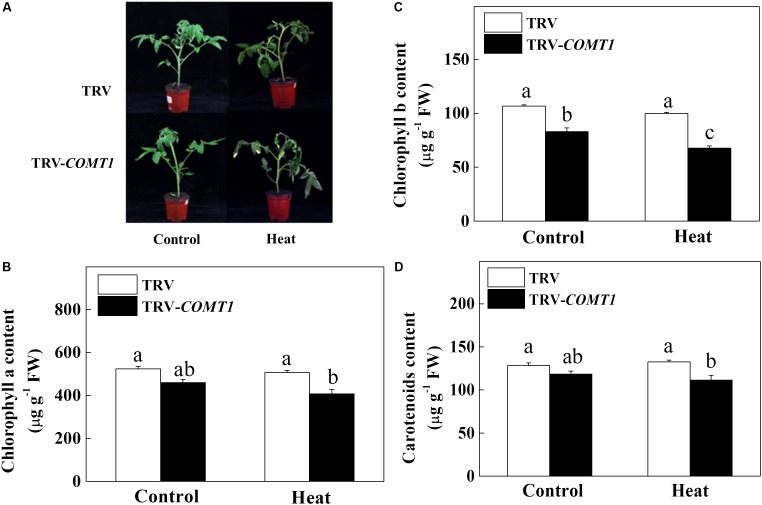
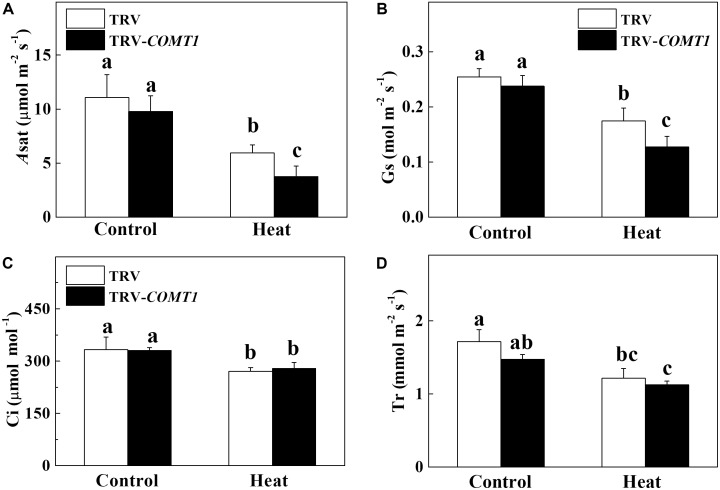
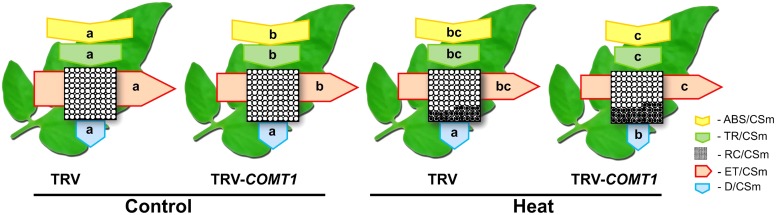
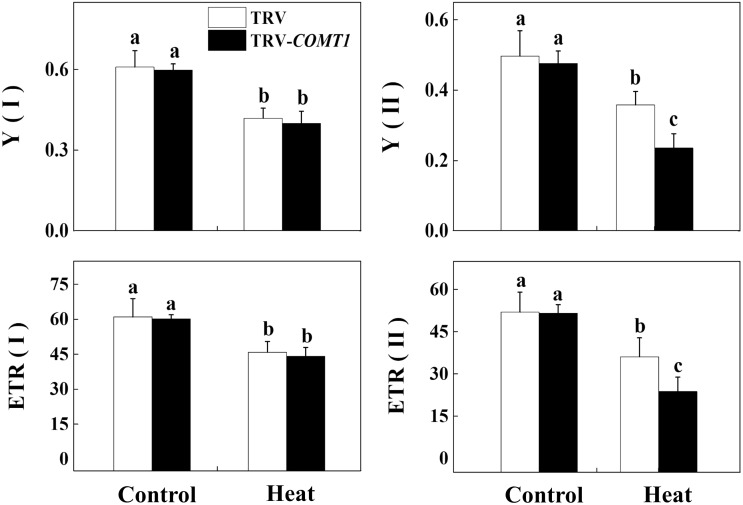
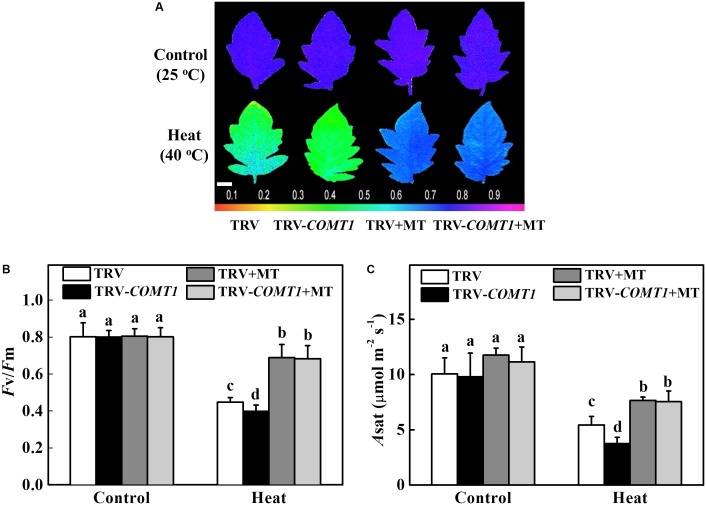
Despite a range of initiatives to reduce global carbon emission, the mean global temperature is increasing due to climate change. Since rising temperatures pose a serious threat of food insecurity, it is important to further explore important biological molecules that can confer thermotolerance to plants. Recently, melatonin has emerged as a universal abiotic stress regulator that can enhance plant tolerance to high temperature. Nonetheless, such regulatory roles of melatonin were unraveled mainly by assessing the effect of exogenous melatonin on plant tolerance to abiotic stress. Here, we generated melatonin deficient tomato plants by silencing of a melatonin biosynthetic gene, CAFFEIC ACID O-METHYLTRANSFERASE 1 (COMT1), to unveil the role of endogenous melatonin in photosynthesis under heat stress. We examined photosynthetic pigment content, leaf gas exchange, and a range of chlorophyll fluorescence parameters. The results showed that silencing of COMT1 aggravated heat stress by inhibiting both the light reactions and the carbon fixation reactions of photosynthesis. The photosynthetic pigment content, light absorption flux, trapped energy flux, energy dissipation, density of active reaction center per photosystem II (PSII) cross-section, the photosynthetic electron transport rate, the maximum photochemical efficiency of PSII photochemistry, and the rate of CO2 assimilation all decreased in COMT1-silenced plants compared with that of non-silenced plants particularly under heat stress. However, exogenous melatonin alleviated heat-induced photosynthetic inhibition in both genotypes, indicating that melatonin is essential for maintaining photosynthetic capacity under stressful conditions. These findings provide genetic evidence on the vital role of melatonin in photosynthesis and thus may have useful implication in horticultural crop management in the face of climate change.
Keywords: JIP test; chlorophyll fluorescence; heat stress; melatonin; photosynthesis; tomato.
Figures
References
-
- Ahammed G. J., Li X., Zhou J., Zhou Y. H., Yu J. Q. (2016). “Role of hormones in plant adaptation to heat stress,” in Plant Hormones Under Challenging Environmental Factors, eds Ahammed G. J., Yu J. Q. (Dordrecht: Springer; ), 1–21.
-
- Brestic M., Zivcak M. (2013). “PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: protocols and applications,” in Molecular Stress Physiology of Plants, eds Rout G. R., Das A. B. (New Delhi: Springer India; ), 87–131. 10.1007/978-81-322-0807-5_4 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources