

Thalidomide promotes degradation of SALL4, a transcription factor implicated in Duane Radial Ray syndrome
- PMID: 30067223
- PMCID: PMC6156078
- DOI: 10.7554/eLife.38430
Thalidomide promotes degradation of SALL4, a transcription factor implicated in Duane Radial Ray syndrome
Abstract
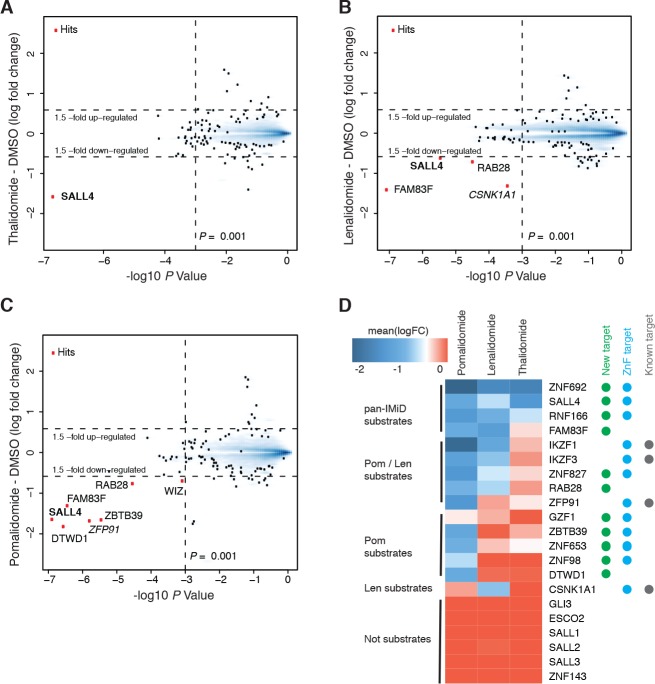
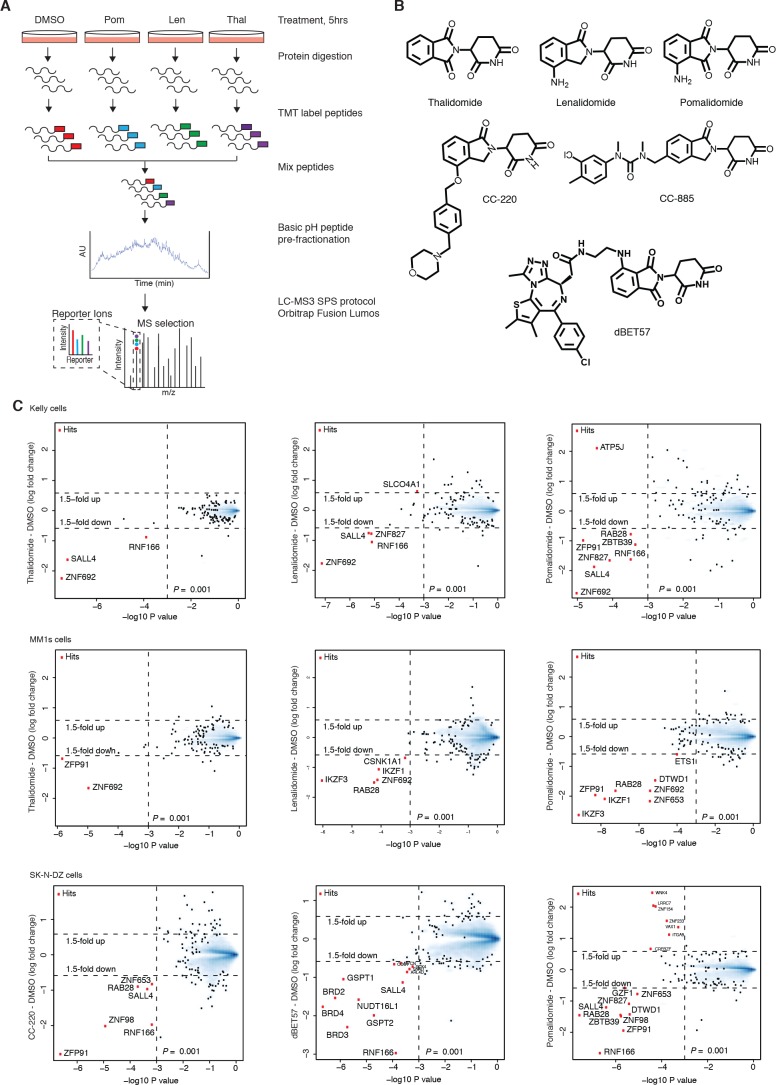
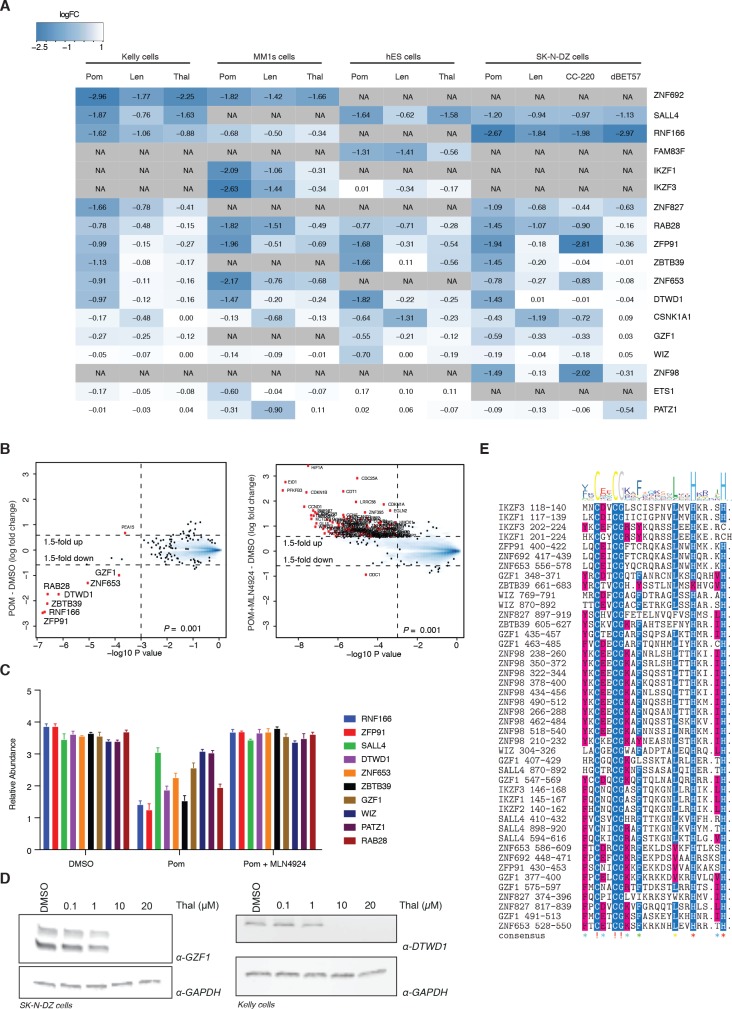
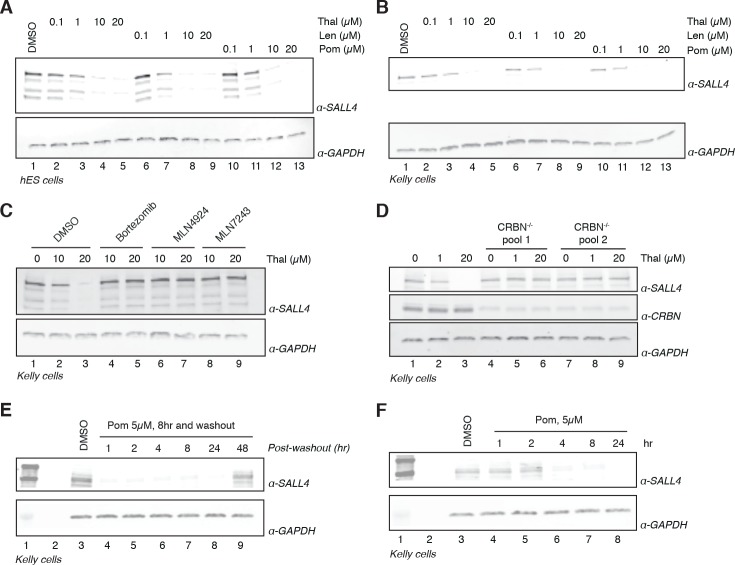
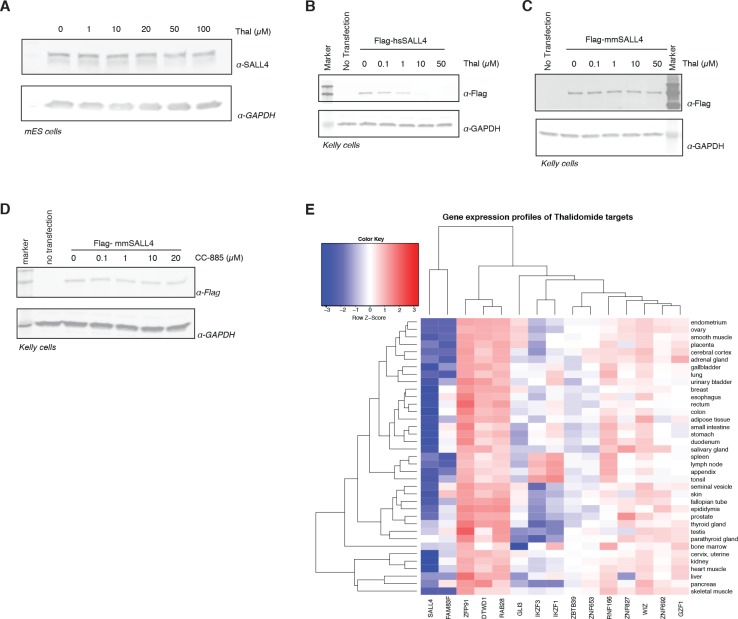
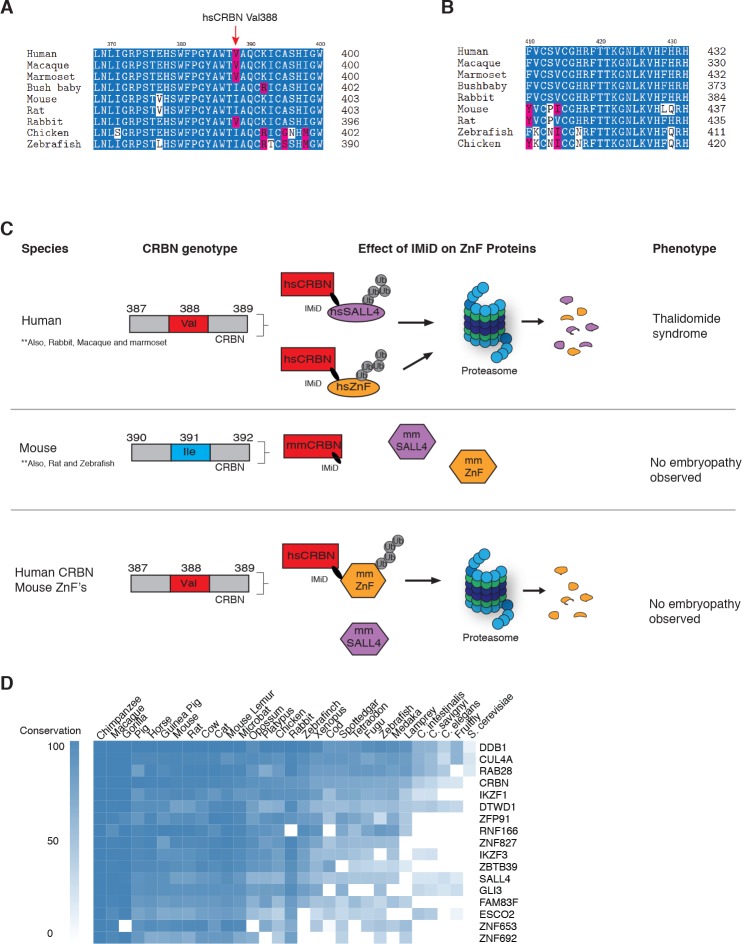
In historical attempts to treat morning sickness, use of the drug thalidomide led to the birth of thousands of children with severe birth defects. Despite their teratogenicity, thalidomide and related IMiD drugs are now a mainstay of cancer treatment; however, the molecular basis underlying the pleiotropic biology and characteristic birth defects remains unknown. Here we show that IMiDs disrupt a broad transcriptional network through induced degradation of several C2H2 zinc finger transcription factors, including SALL4, a member of the spalt-like family of developmental transcription factors. Strikingly, heterozygous loss of function mutations in SALL4 result in a human developmental condition that phenocopies thalidomide-induced birth defects such as absence of thumbs, phocomelia, defects in ear and eye development, and congenital heart disease. We find that thalidomide induces degradation of SALL4 exclusively in humans, primates, and rabbits, but not in rodents or fish, providing a mechanistic link for the species-specific pathogenesis of thalidomide syndrome.
Keywords: biochemistry; chemical biology; developmental biology; human; teratogenicity; transcription factors; ubiquitin.
© 2018, Donovan et al.
Conflict of interest statement
KD, JA, RN, JY, EF, BB, BE No competing interests declared, EF is a member of the scientific advisory board of C4 Therapeutics.
Figures
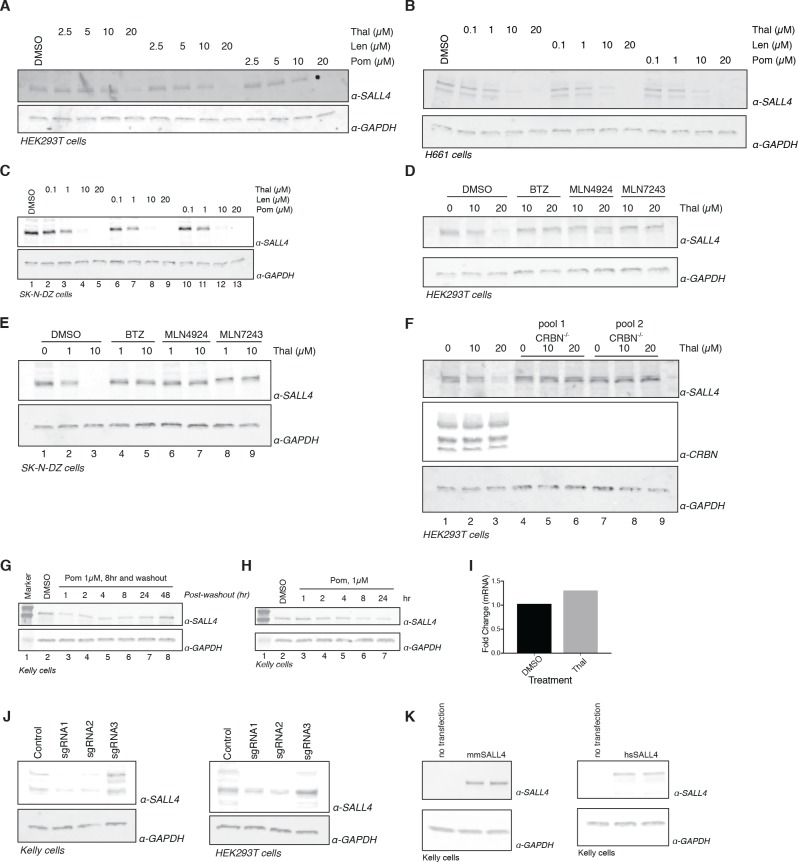
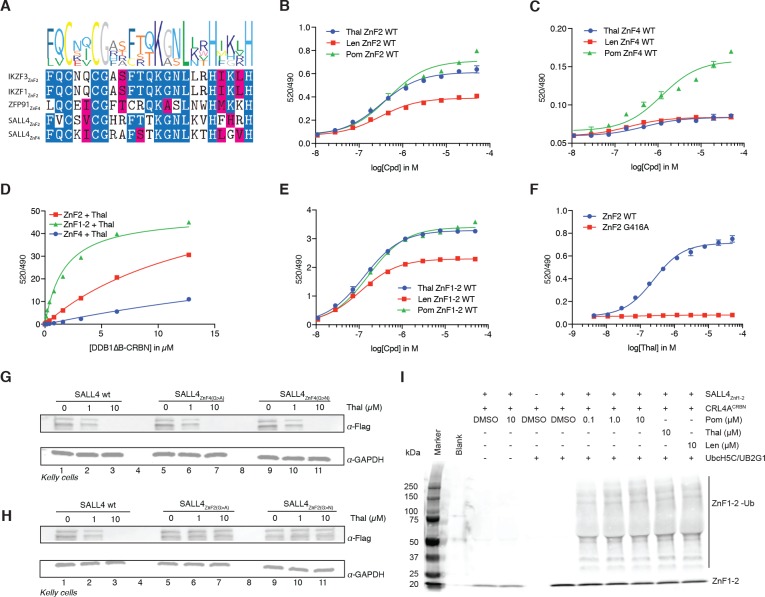
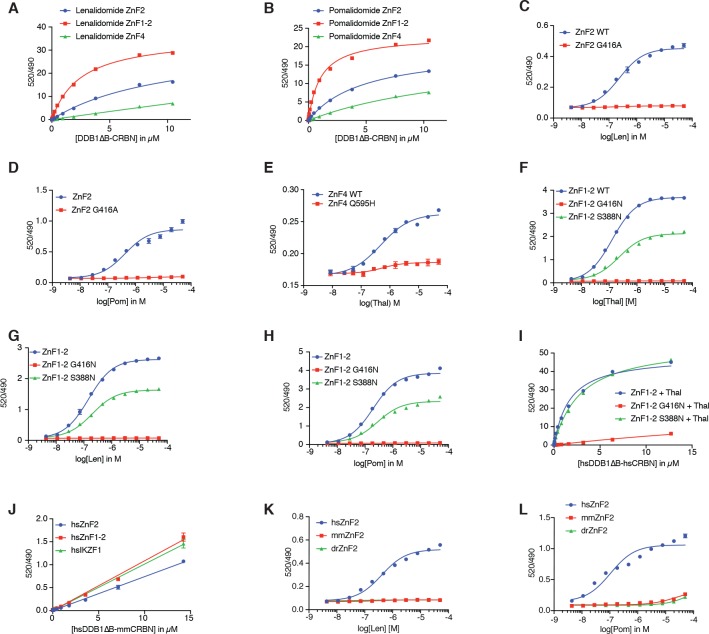
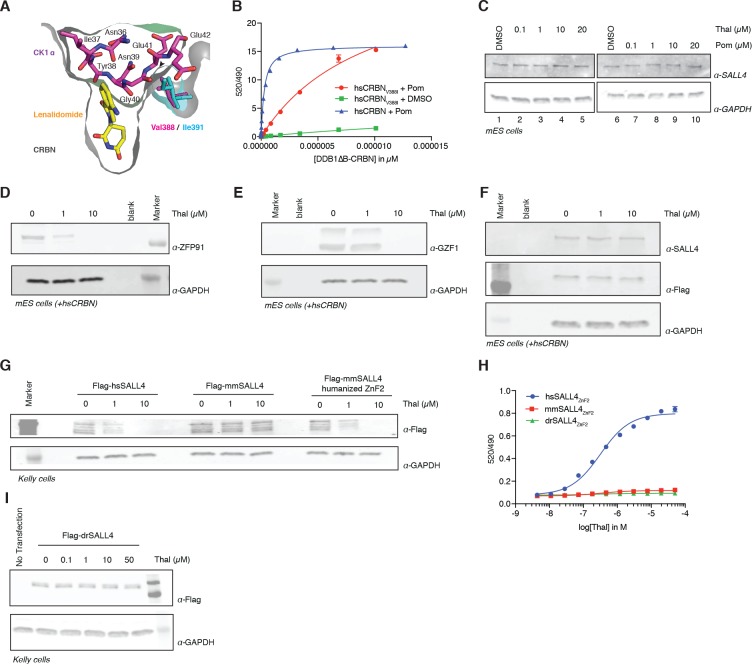
Comment in
-
Unlocking a dark past.Elife. 2018 Sep 25;7:e41002. doi: 10.7554/eLife.41002. Elife. 2018. PMID: 30252647 Free PMC article.
References
-
- Cavadini S, Fischer ES, Bunker RD, Potenza A, Lingaraju GM, Goldie KN, Mohamed WI, Faty M, Petzold G, Beckwith RE, Tichkule RB, Hassiepen U, Abdulrahman W, Pantelic RS, Matsumoto S, Sugasawa K, Stahlberg H, Thomä NH. Cullin-RING ubiquitin E3 ligase regulation by the COP9 signalosome. Nature. 2016;531:598–603. doi: 10.1038/nature17416. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
