

Ki67 is a Graded Rather than a Binary Marker of Proliferation versus Quiescence
- PMID: 30067968
- PMCID: PMC6108547
- DOI: 10.1016/j.celrep.2018.06.110
Ki67 is a Graded Rather than a Binary Marker of Proliferation versus Quiescence
Abstract
Ki67 staining is widely used as a proliferation indicator in the clinic, despite poor understanding of this protein's function or dynamics. Here, we track Ki67 levels under endogenous control in single cells over time and find that Ki67 accumulation occurs only during S, G2, and M phases. Ki67 is degraded continuously in G1 and G0 phases, regardless of the cause of entry into G0/quiescence. Consequently, the level of Ki67 during G0 and G1 in individual cells is highly heterogeneous and depends on how long an individual cell has spent in G0. Thus, Ki67 is a graded rather than a binary marker both for cell-cycle progression and time since entry into quiescence.
Keywords: CDK2 activity; Ki67; heterogeneity; quiescence; single-cell tracking; time-lapse imaging.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
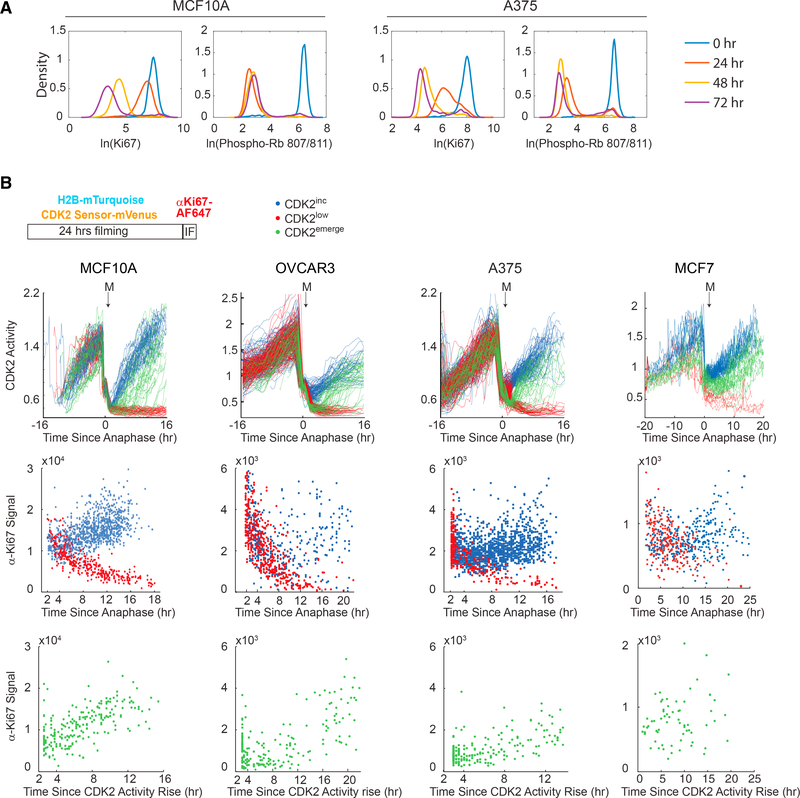
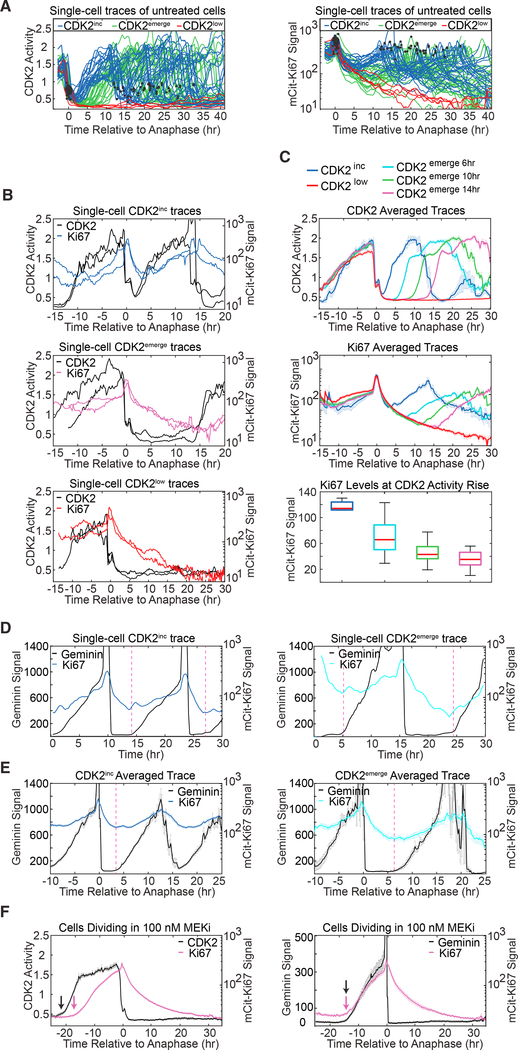
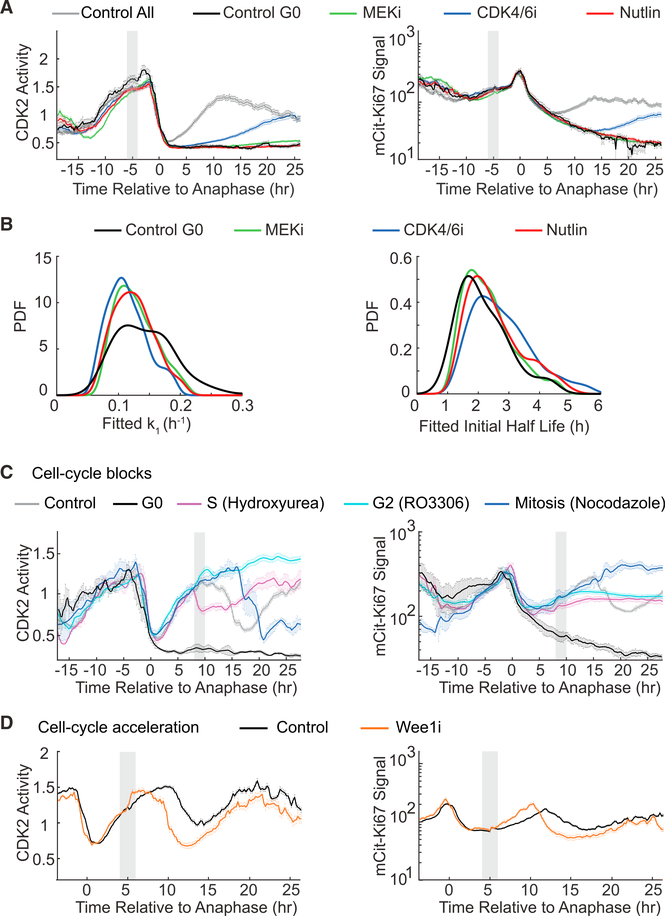
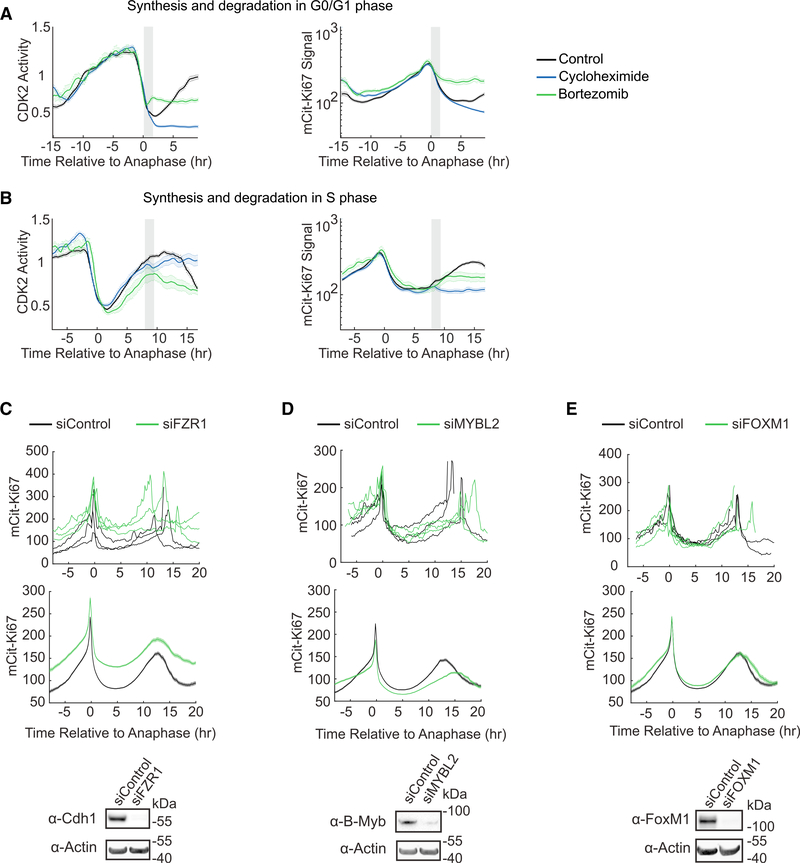
Comment in
-
Ki-67: More Hidden behind a 'Classic Proliferation Marker'.Trends Biochem Sci. 2018 Oct;43(10):747-748. doi: 10.1016/j.tibs.2018.08.004. Epub 2018 Aug 18. Trends Biochem Sci. 2018. PMID: 30131191
References
-
- Dowsett M, and Dunbier AK (2008). Emerging biomarkers and new understanding of traditional markers in personalized therapy for breast cancer. Clin. Cancer Res 14, 8019–8026. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
