

Timed Regulation of 3BP2 Induction Is Critical for Sustaining CD8+ T Cell Expansion and Differentiation
- PMID: 30067970
- PMCID: PMC6701191
- DOI: 10.1016/j.celrep.2018.06.075
Timed Regulation of 3BP2 Induction Is Critical for Sustaining CD8+ T Cell Expansion and Differentiation
Abstract
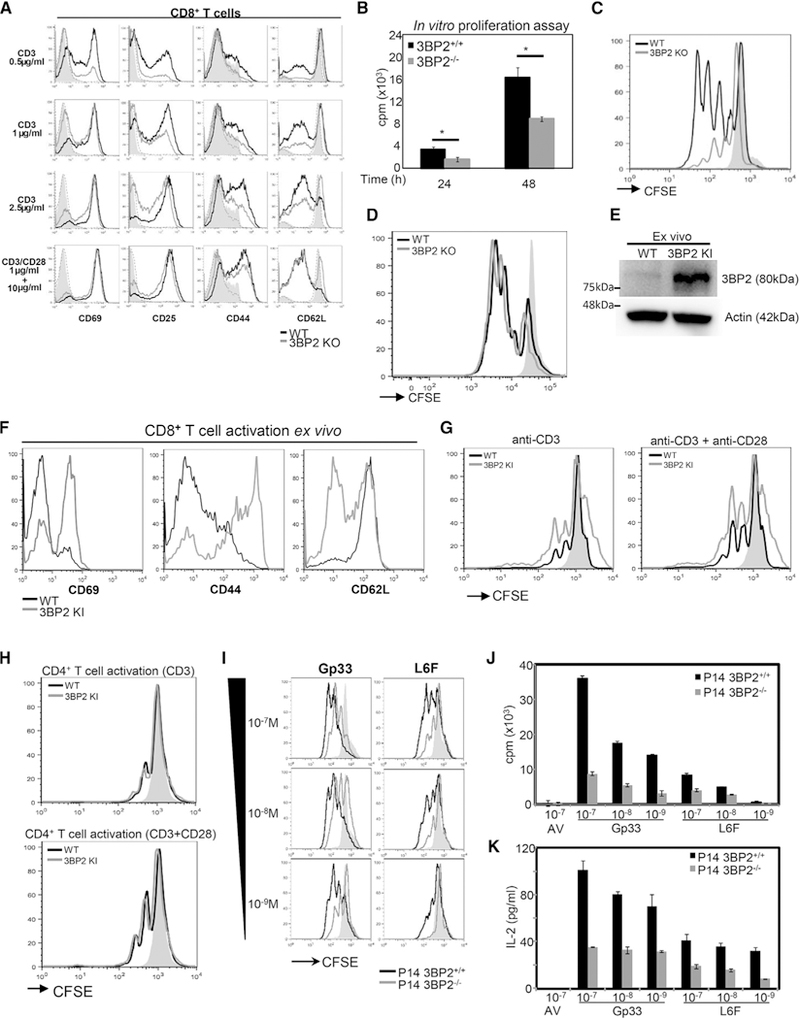
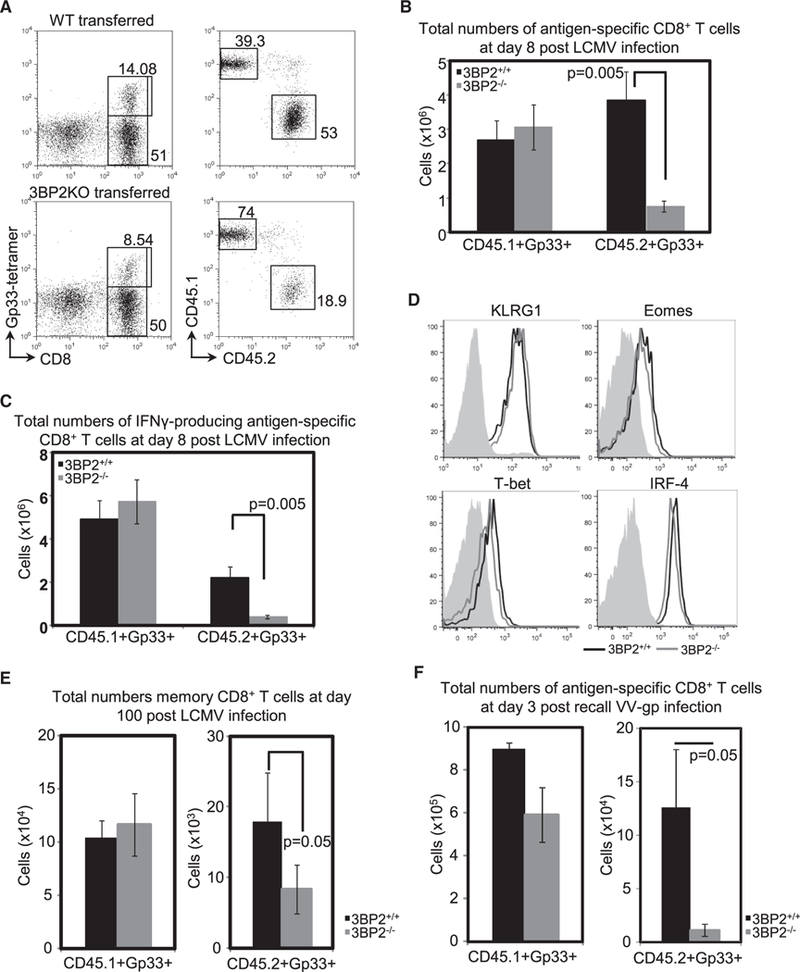
Successful anti-viral response requires the sustained activation and expansion of CD8+ T cells for periods that far exceed the time limit of physical T cell interaction with antigen-presenting cells (APCs). The expanding CD8+ T cell pool generates the effector and memory cell populations that provide viral clearance and long-term immunity, respectively. Here, we demonstrate that 3BP2 is recruited in cytoplasmic microclusters and nucleates a signaling complex that facilitates MHC:peptide-independent activation of signaling pathways downstream of the TCR. We show that induction of the adaptor molecule 3BP2 is a sensor of TCR signal strength and is critical for sustaining CD8+ T cell proliferation and regulating effector and memory differentiation.
Keywords: 3BP2; CD8(+) T cells; TCR signaling; differentiation; microclusters; proliferation.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures





References
-
- Bunnell SC, Barr VA, Fuller CL, and Samelson LE (2003). High-resolu-tion multi color imaging of dynamic signaling complexes in T cells stimulated by planar substrates. Sci. STKE 2003, PL8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
