

Small molecule-based targeting of TTD-A dimerization to control TFIIH transcriptional activity represents a potential strategy for anticancer therapy
- PMID: 30068551
- PMCID: PMC6166727
- DOI: 10.1074/jbc.RA118.003444
Small molecule-based targeting of TTD-A dimerization to control TFIIH transcriptional activity represents a potential strategy for anticancer therapy
Abstract
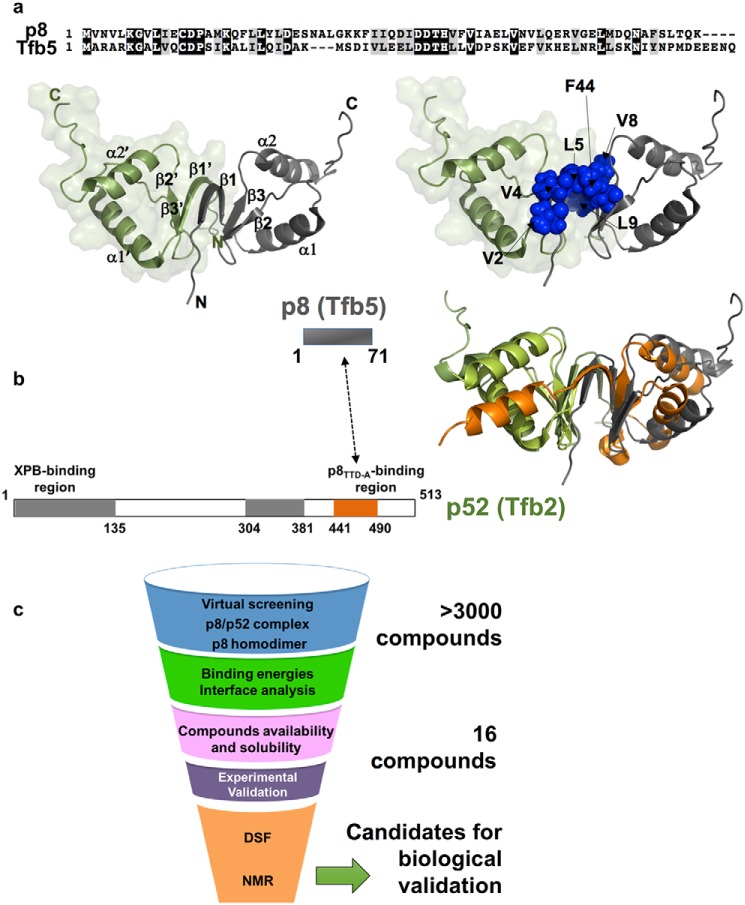
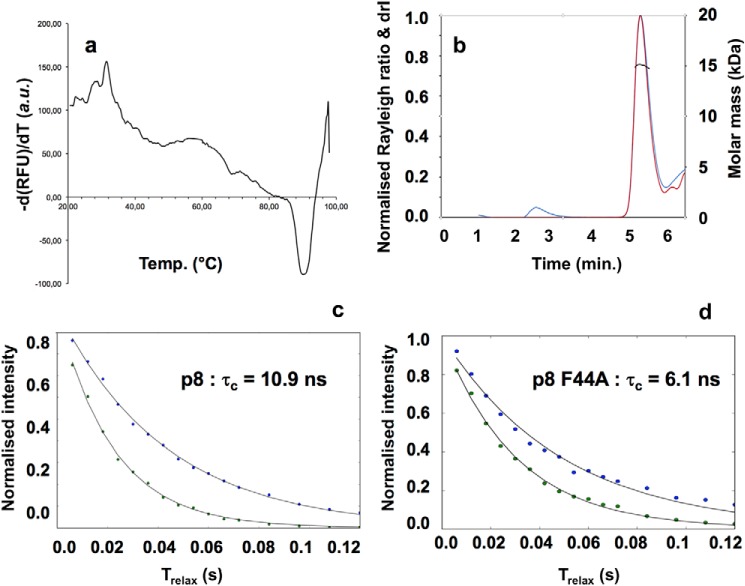
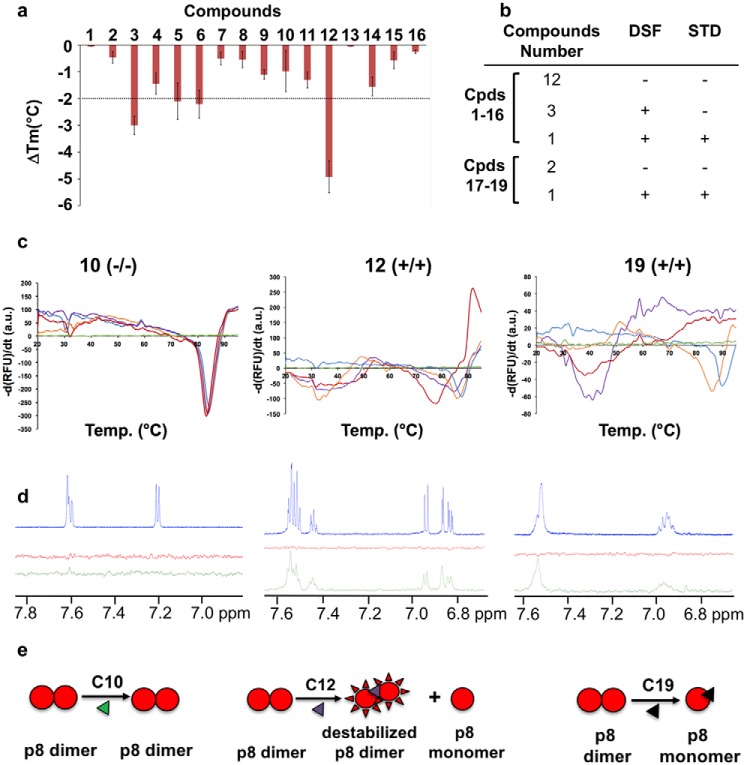
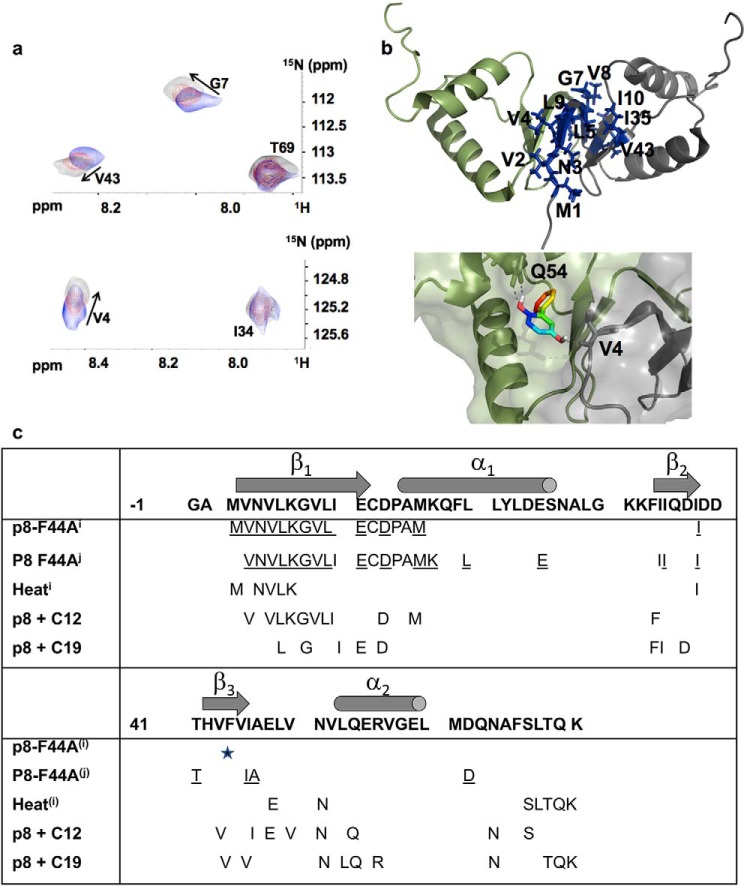
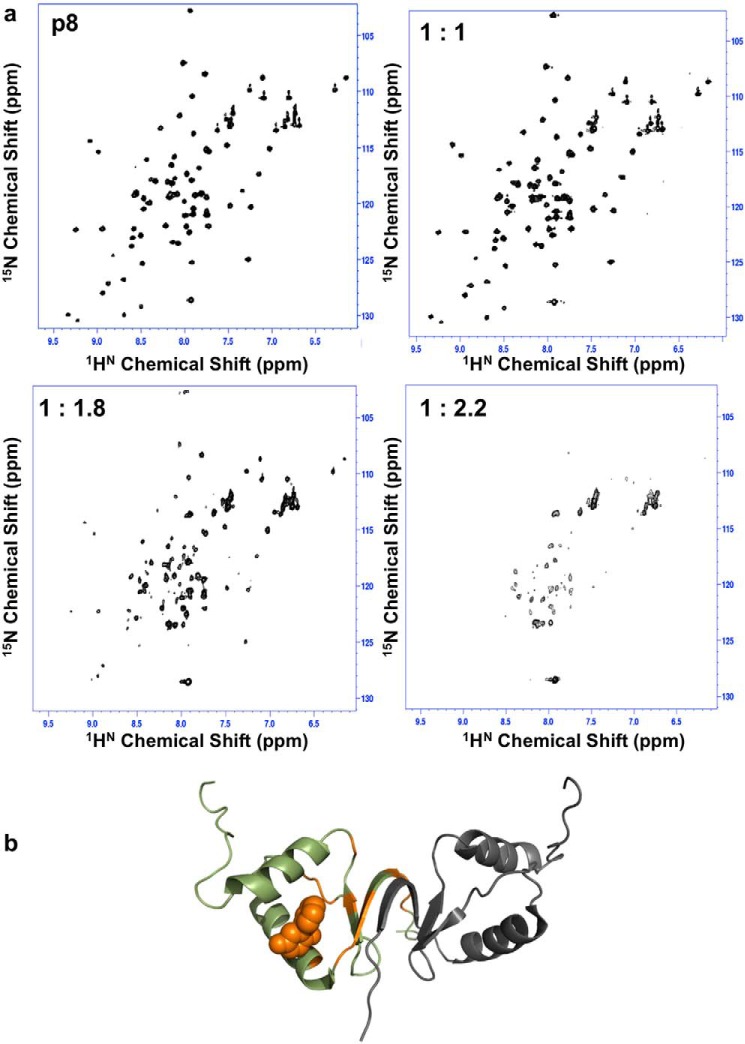
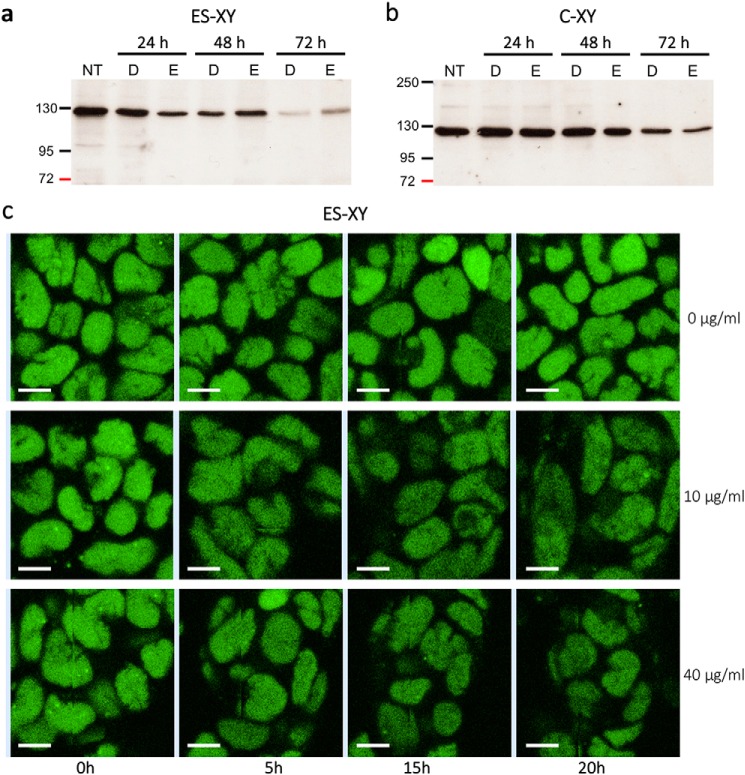
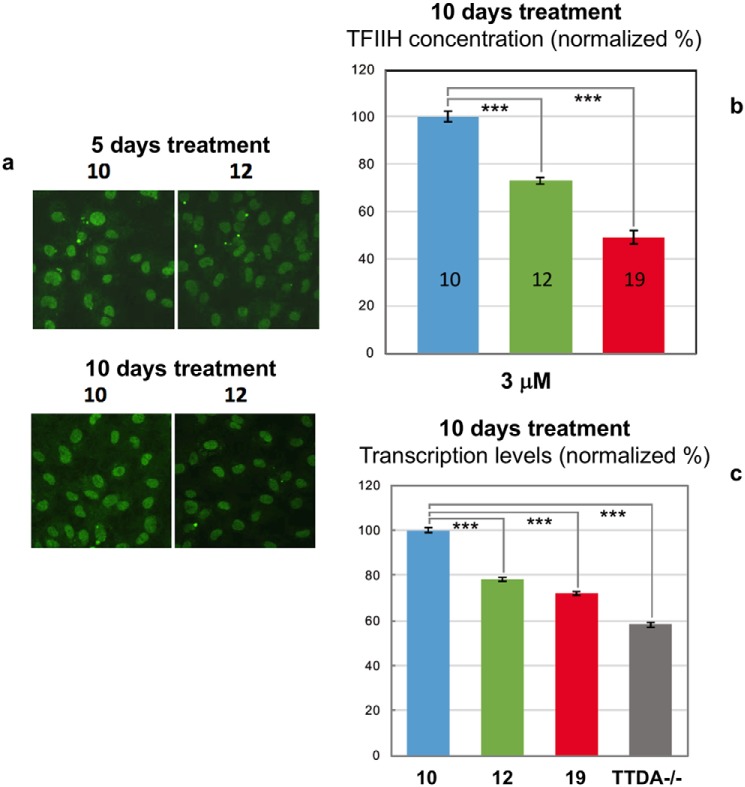
The human transcription factor TFIIH is a large complex composed of 10 subunits that form an intricate network of protein-protein interactions critical for regulating its transcriptional and DNA repair activities. The trichothiodystrophy group A protein (TTD-A or p8) is the smallest TFIIH subunit, shuttling between a free and a TFIIH-bound state. Its dimerization properties allow it to shift from a homodimeric state, in the absence of a functional partner, to a heterodimeric structure, enabling dynamic binding to TFIIH. Recruitment of p8 at TFIIH stabilizes the overall architecture of the complex, whereas p8's absence reduces its cellular steady-state concentration and consequently decreases basal transcription, highlighting that p8 dimerization may be an attractive target for down-regulating transcription in cancer cells. Here, using a combination of molecular dynamics simulations to study p8 conformational stability and a >3000-member library of chemical fragments, we identified small-molecule compounds that bind to the dimerization interface of p8 and provoke its destabilization, as assessed by biophysical studies. Using quantitative imaging of TFIIH in living mouse cells, we found that these molecules reduce the intracellular concentration of TFIIH and its transcriptional activity to levels similar to that observed in individuals with trichothiodystrophy owing to mutated TTD-A Our results provide a proof of concept of fragment-based drug discovery, demonstrating the utility of small molecules for targeting p8 dimerization to modulate the transcriptional machinery, an approach that may help inform further development in anticancer therapies.
Keywords: DNA transcription; GTF2H5; STD-NMR; TFIIH; cancer; drug screening; nuclear magnetic resonance (NMR); protein-protein interaction; quantitative imaging; transcriptional regulation.
© 2018 Gervais et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Luo J., Cimermancic P., Viswanath S., Ebmeier C. C., Kim B., Dehecq M., Raman V., Greenberg C. H., Pellarin R., Sali A., Taatjes D. J., Hahn S., and Ranish J. (2015) Architecture of the human and yeast general transcription and DNA repair factor TFIIH. Mol. Cell 59, 794–806 10.1016/j.molcel.2015.07.016 - DOI - PMC - PubMed
-
- Giglia-Mari G., Coin F., Ranish J. A., Hoogstraten D., Theil A., Wijgers N., Jaspers N. G. J., Raams A., Argentini M., van der Spek P. J., Botta E., Stefanini M., Egly J. M., Aebersold R., Hoeijmakers J. H. J., and Vermeulen W. (2004) A new, tenth subunit of TFIIH is responsible for the DNA repair syndrome trichothiodystrophy group A. Nat. Genet. 36, 714–719 10.1038/ng1387 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
