

ERK Signaling Controls Innate-like CD8+ T Cell Differentiation via the ELK4 (SAP-1) and ELK1 Transcription Factors
- PMID: 30068599
- PMCID: PMC6121213
- DOI: 10.4049/jimmunol.1800704
ERK Signaling Controls Innate-like CD8+ T Cell Differentiation via the ELK4 (SAP-1) and ELK1 Transcription Factors
Abstract
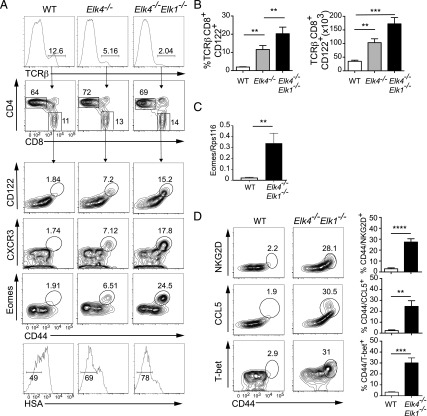
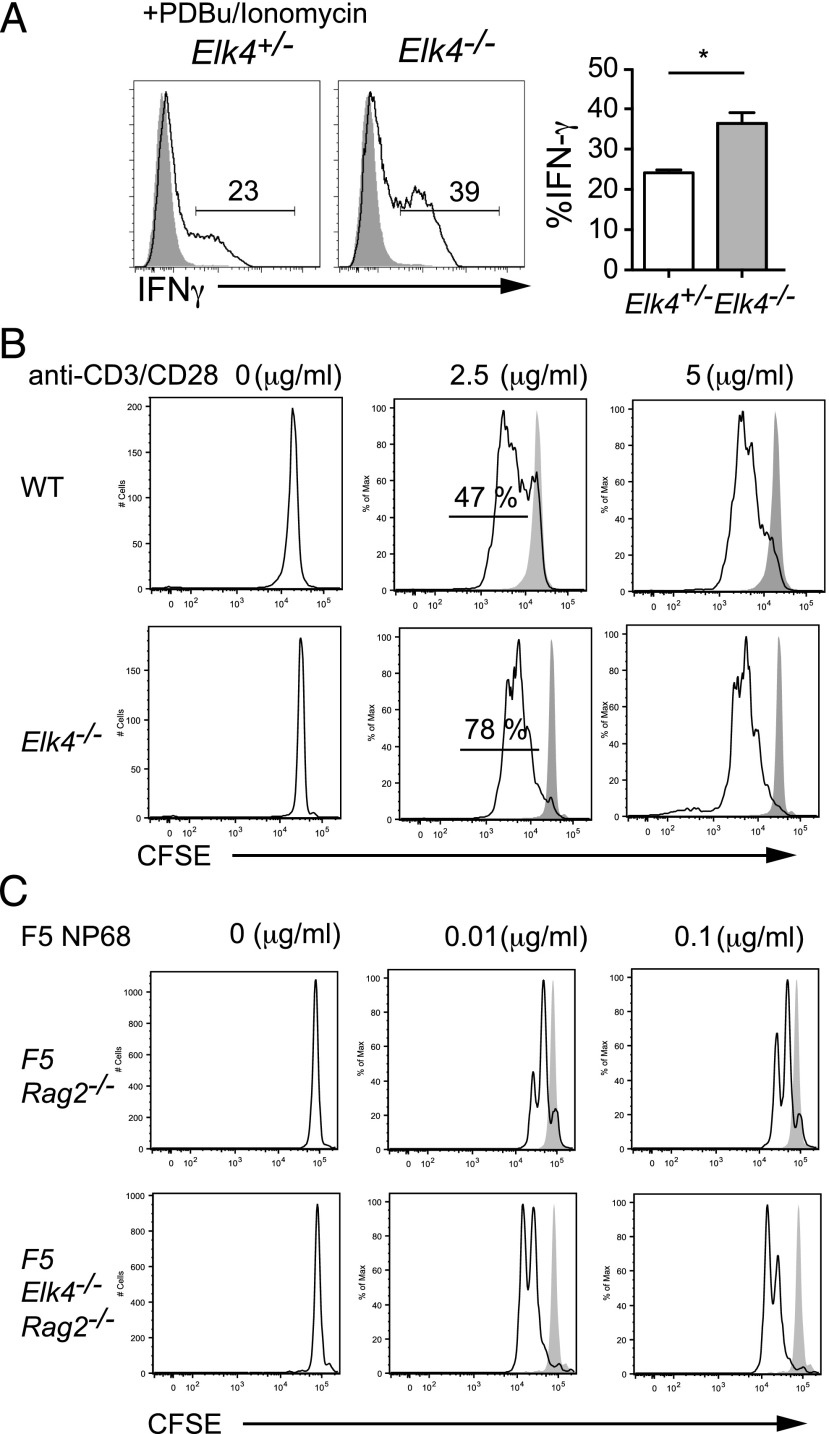
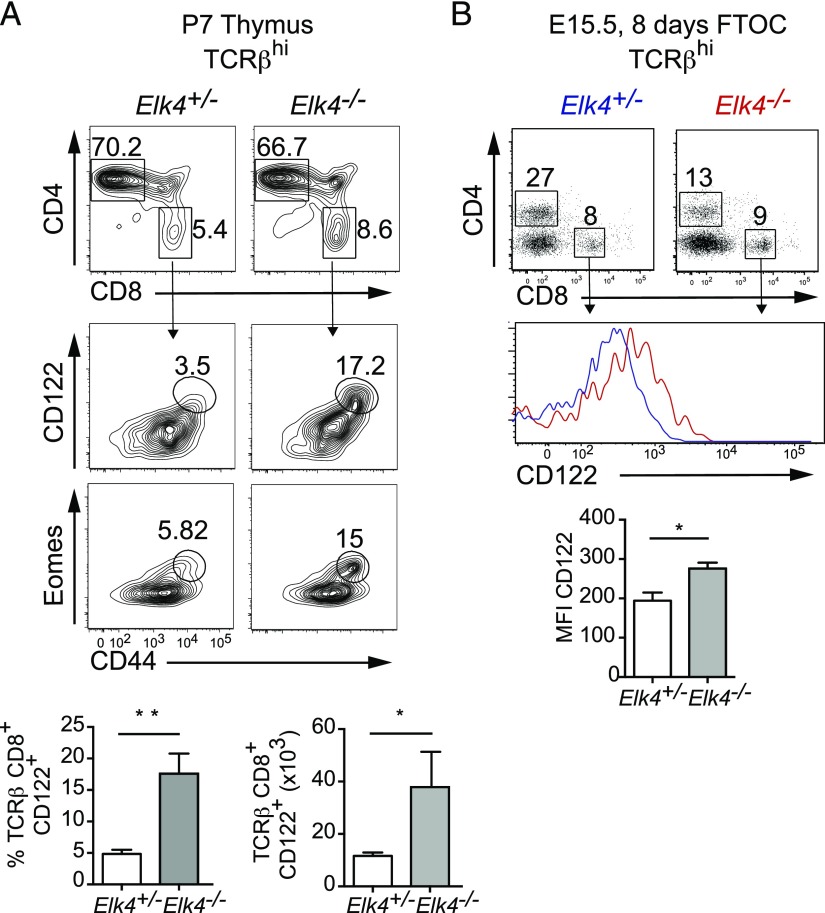
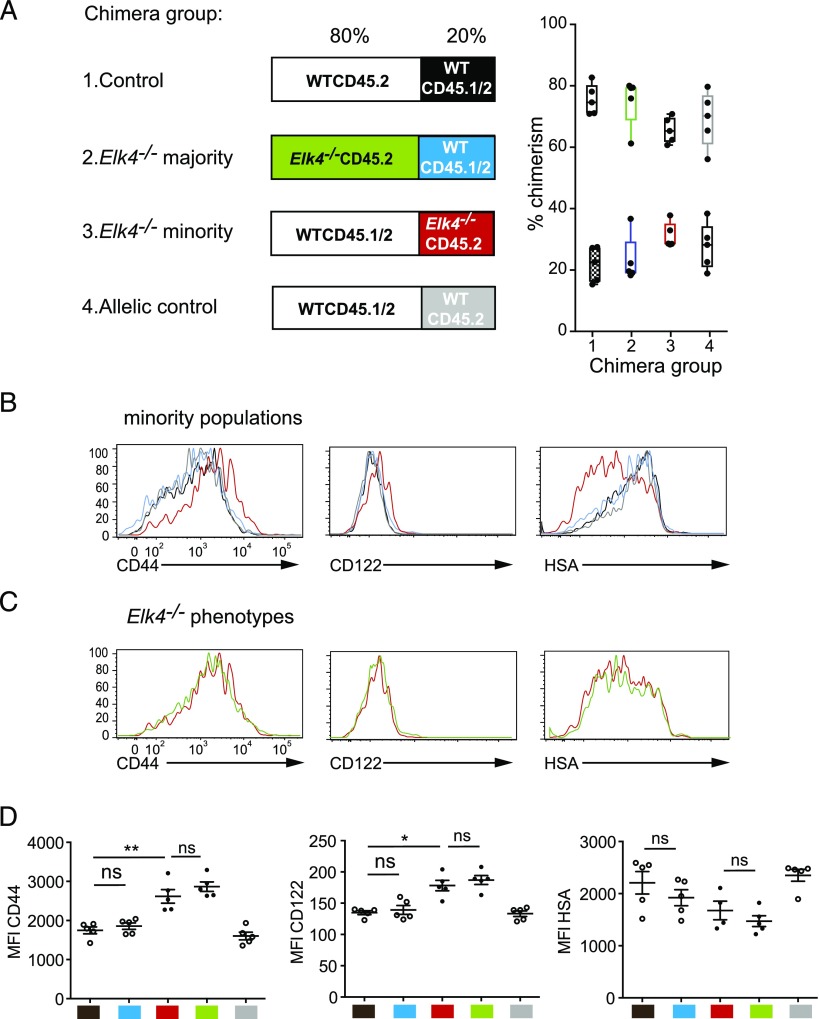
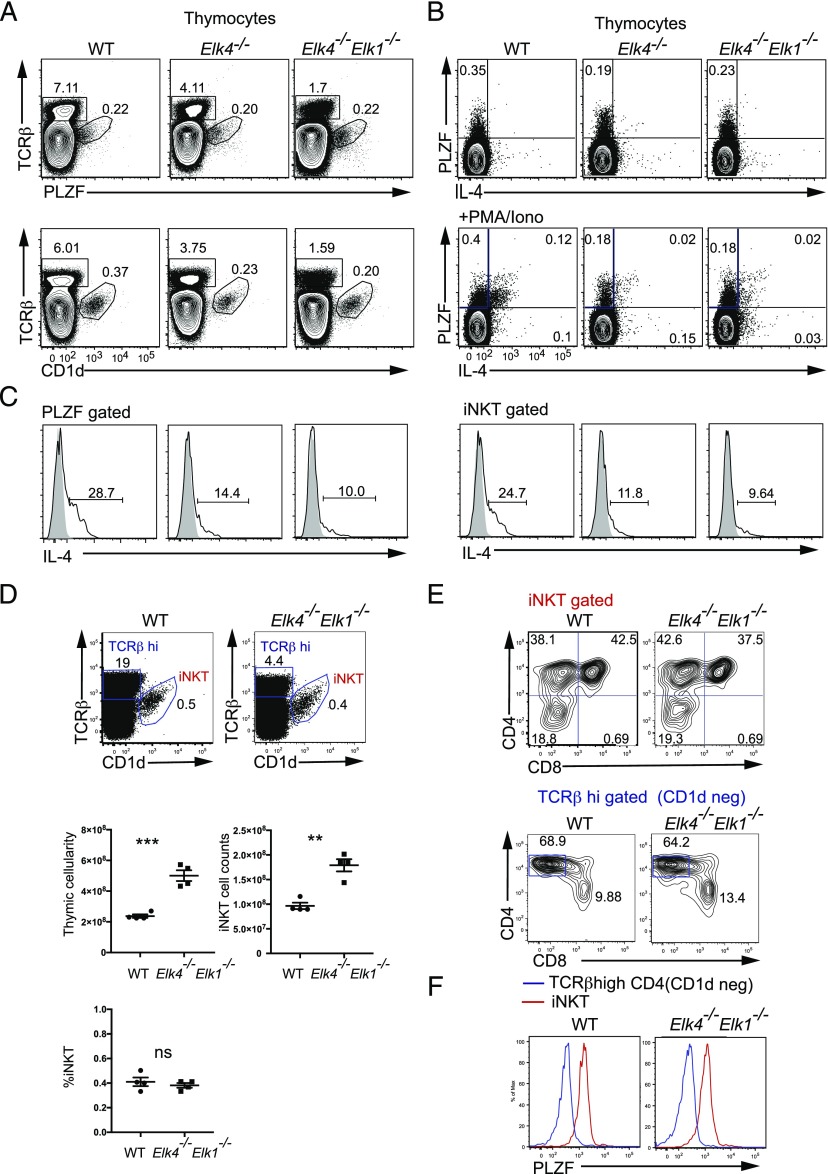
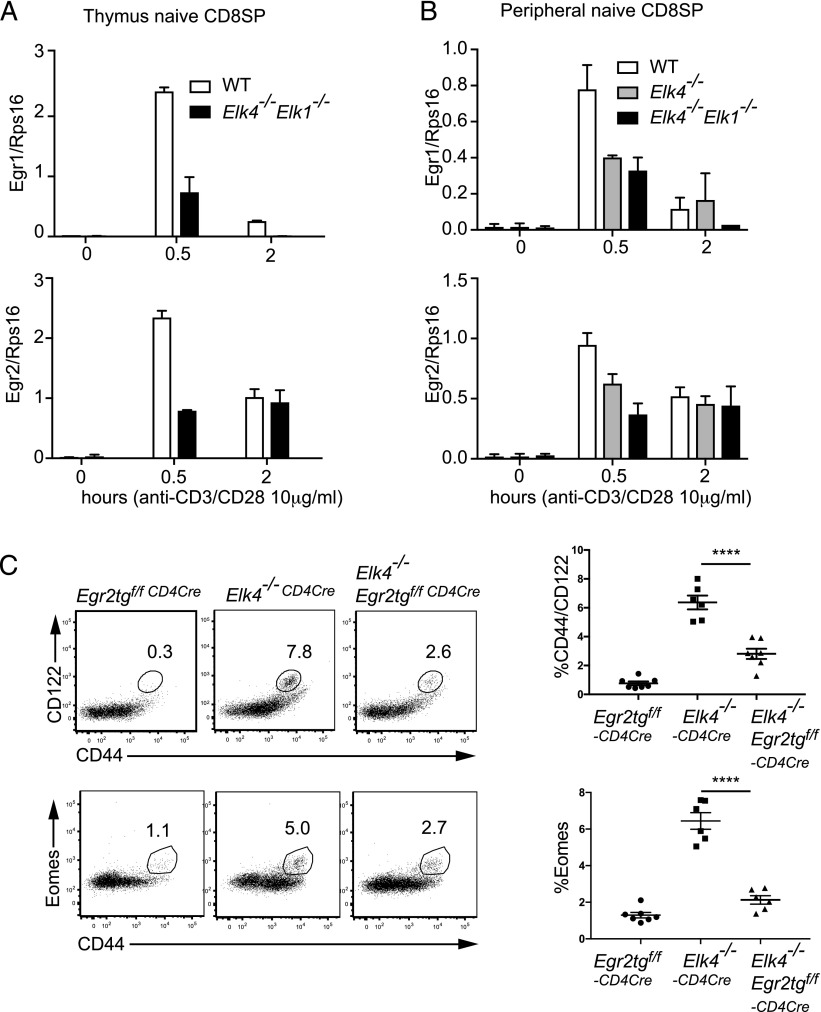
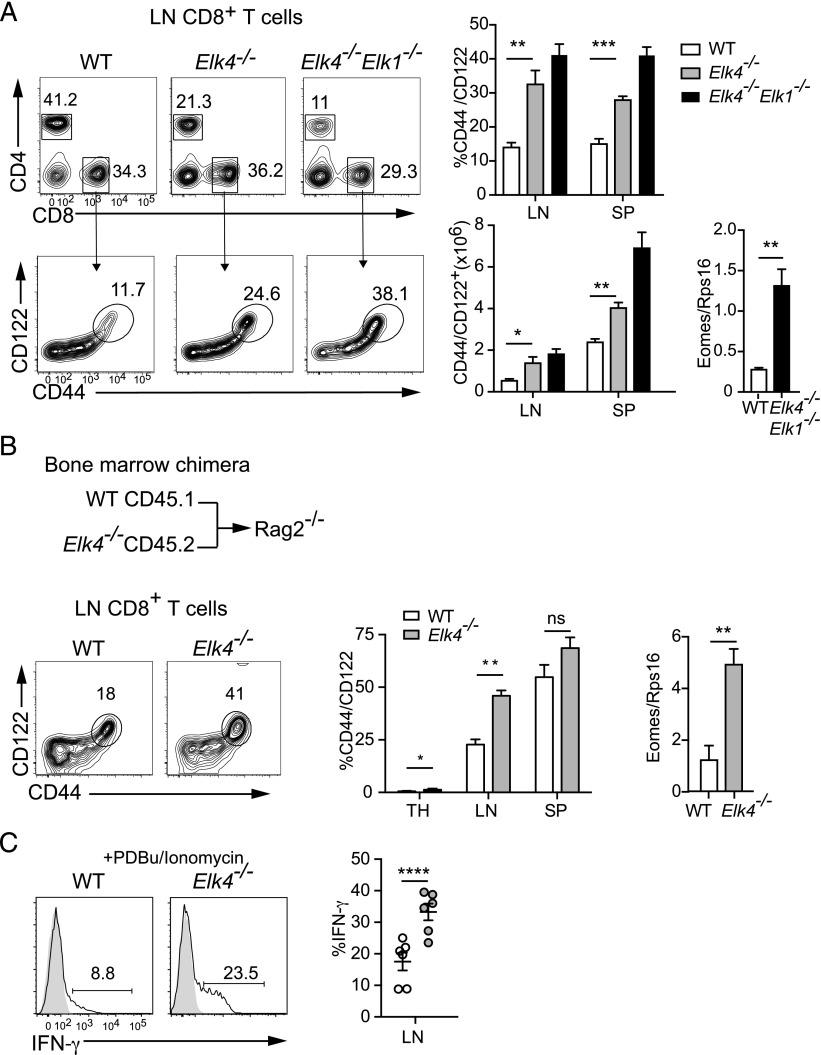
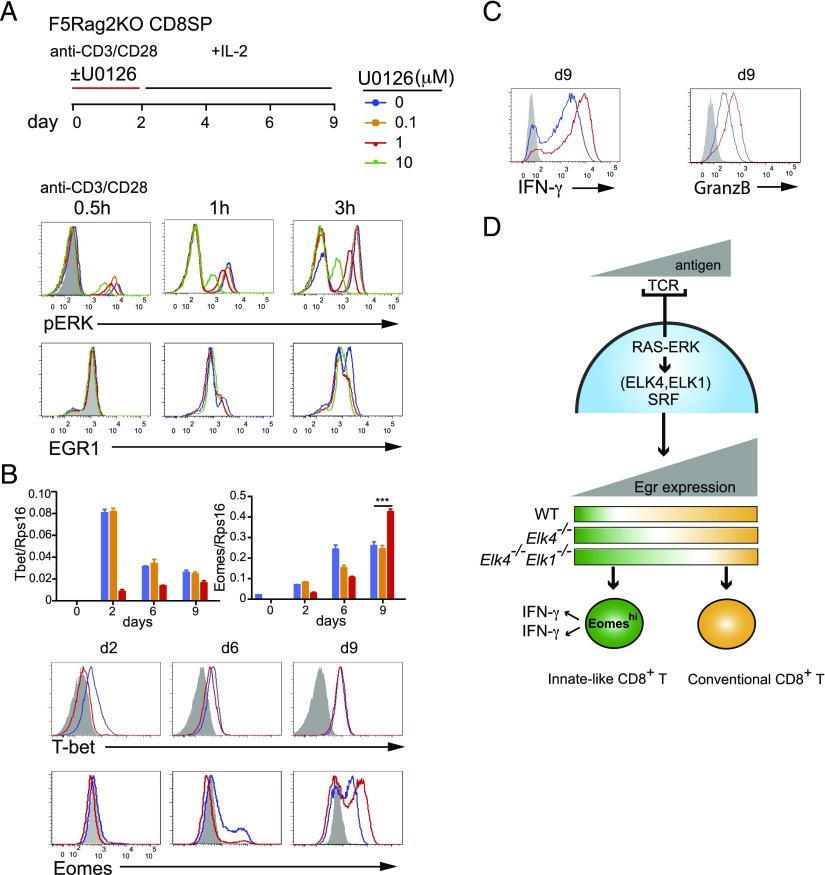
In mouse thymocyte development, signaling by the TCR through the ERK pathway is required for positive selection of conventional naive T cells. The Ets transcription factor ELK4 (SAP-1), an ERK-regulated cofactor of the SRF transcription factor, plays an important role in positive selection by activating immediate-early genes such as the Egr transcription factor family. The role of ELK4-SRF signaling in development of other T cell types dependent on ERK signaling has been unclear. In this article, we show that ELK4, and its close relative ELK1, act cell autonomously in the thymus to control the generation of innate-like αβ CD8+ T cells with memory-like characteristics. Mice lacking ELK4 and ELK1 develop increased numbers of innate-like αβ CD8+ T cells, which populate the periphery. These cells develop cell autonomously rather than through expansion of PLZF+ thymocytes and concomitantly increased IL-4 signaling. Their development is associated with reduced TCR-mediated activation of ELK4-SRF target genes and can be partially suppressed by overexpression of the ELK4-SRF target gene EGR2. Consistent with this, partial inhibition of ERK signaling in peripheral CD8+T cells promotes the generation of cells with innate-like characteristics. These data establish that low-level ERK signaling through ELK4 (and ELK1) promotes innate-like αβ CD8+ T cell differentiation, tuning conventional versus innate-like development.
Copyright © 2018 The Authors.
Figures
References
-
- Gascoigne N. R., Rybakin V., Acuto O., Brzostek J. 2016. TCR signal strength and T cell development. Annu. Rev. Cell Dev. Biol. 32: 327–348. - PubMed
-
- Starr T. K., Jameson S. C., Hogquist K. A. 2003. Positive and negative selection of T cells. Annu. Rev. Immunol. 21: 139–176. - PubMed
-
- Fischer A. M., Katayama C. D., Pagès G., Pouysségur J., Hedrick S. M. 2005. The role of erk1 and erk2 in multiple stages of T cell development. Immunity 23: 431–443. - PubMed
-
- Berg L. J. 2007. Signalling through TEC kinases regulates conventional versus innate CD8(+) T-cell development. Nat. Rev. Immunol. 7: 479–485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
