

In silico discovery of small molecules that inhibit RfaH recruitment to RNA polymerase
- PMID: 30069925
- PMCID: PMC6595482
- DOI: 10.1111/mmi.14093
In silico discovery of small molecules that inhibit RfaH recruitment to RNA polymerase
Abstract
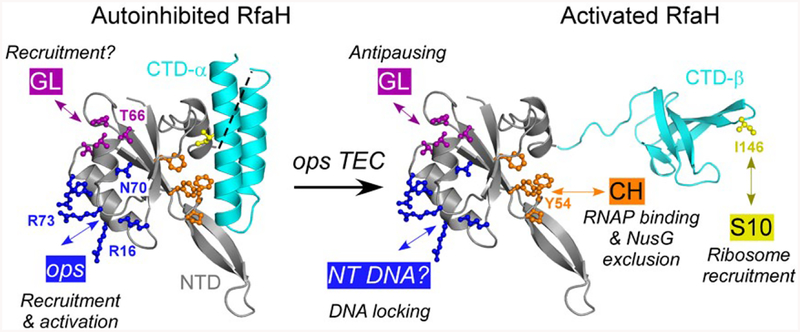
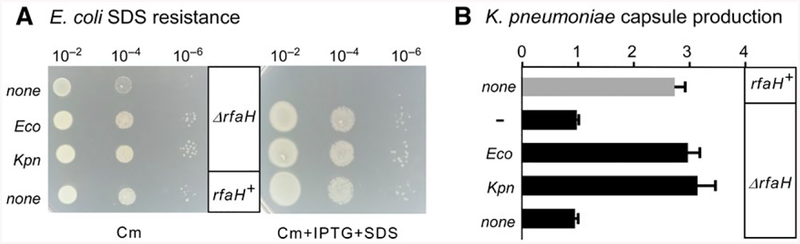
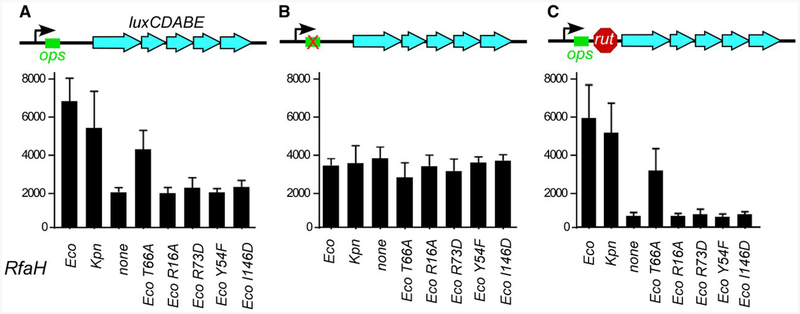
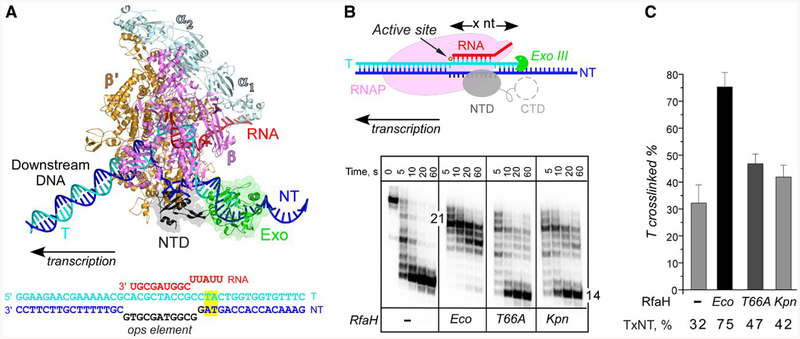
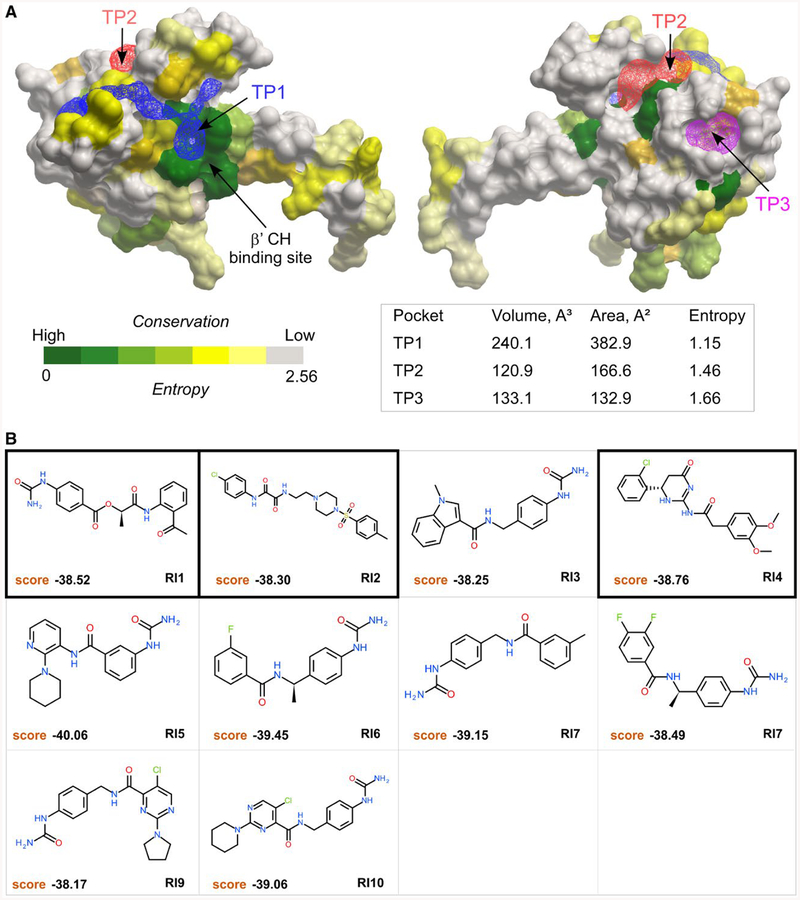
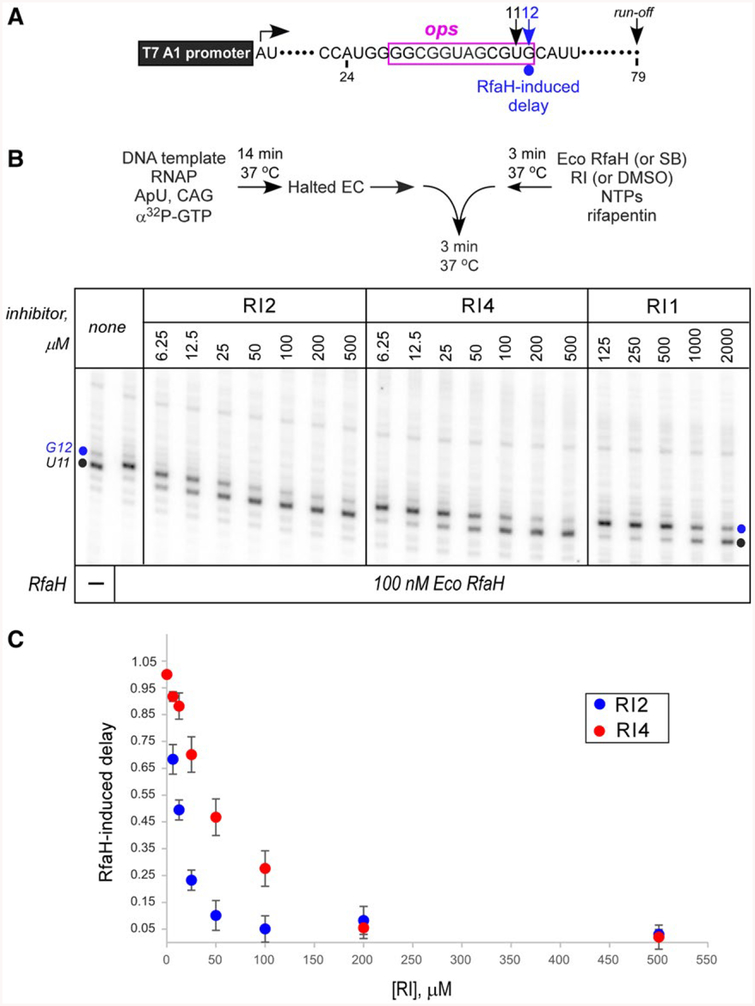
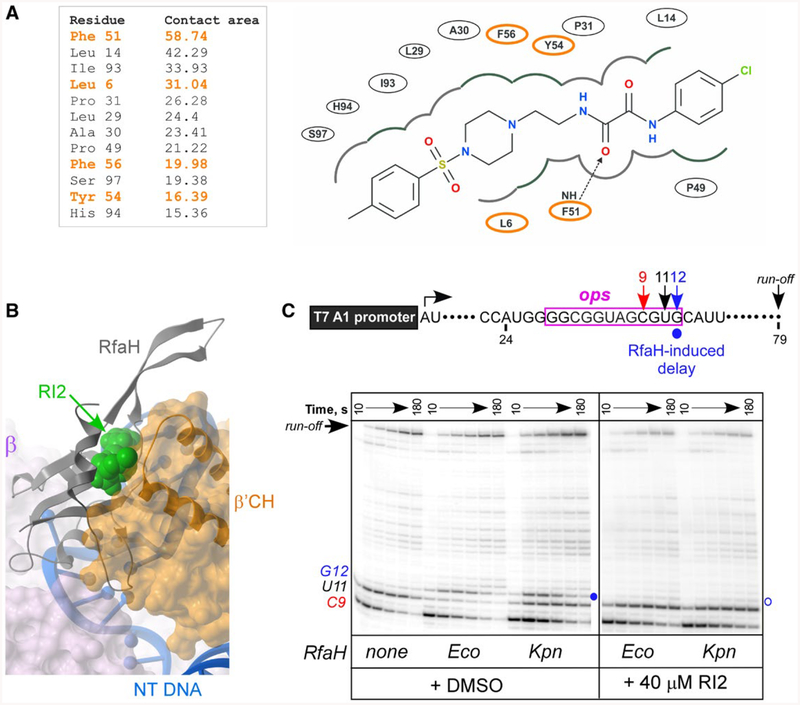
RfaH is required for virulence in several Gram-negative pathogens including Escherichia coli and Klebsiella pneumoniae. Through direct interactions with RNA polymerase (RNAP) and ribosome, RfaH activates the expression of capsule, cell wall and pilus biosynthesis operons by reducing transcription termination and activating translation. While E. coli RfaH has been extensively studied using structural and biochemical approaches, limited data are available for other RfaH homologs. Here we set out to identify small molecule inhibitors of E. coli and K. pneumoniae RfaHs. Results of biochemical and functional assays show that these proteins act similarly, with a notable difference between their interactions with the RNAP β subunit gate loop. We focused on high-affinity RfaH interactions with the RNAP β' subunit clamp helices as a shared target for inhibition. Among the top 10 leads identified by in silico docking using ZINC database, 3 ligands were able to inhibit E. coli RfaH recruitment in vitro. The most potent lead was active against both E. coli and K. pneumoniae RfaHs in vitro. Our results demonstrate the feasibility of identifying RfaH inhibitors using in silico docking and pave the way for rational design of antivirulence therapeutics against antibiotic-resistant pathogens.
© 2018 John Wiley & Sons Ltd.
Conflict of interest statement
Conflict of interest
The Authors declare no conflicts of interest.
Figures
References
-
- Abagyan R, Raush E, Totrov M and Orry A (2017) ICM Manual v3.8–6. San Diego, CA: Molsoft LCC.
-
- Abagyan R, Totrov M and Kuznetsov D (1994) ICM – a new method for protein modeling and design: applications to docking and structure prediction from the distorted native conformation. Journal of Computational Chemistry, 15, 488–506.
-
- An J, Totrov M and Abagyan R (2005) Pocketome via comprehensive identification and classification of ligand binding envelopes. Molecular & Cellular Proteomics, 4, 752–761. - PubMed
-
- Artsimovitch I and Landick R (2002) The transcriptional regulator RfaH stimulates RNA chain synthesis after recruitment to elongation complexes by the exposed non-template DNA strand. Cell, 109, 193–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
