

Roles of distal aspartate and arginine of B-class dye-decolorizing peroxidase in heterolytic hydrogen peroxide cleavage
- PMID: 30072383
- PMCID: PMC6153280
- DOI: 10.1074/jbc.RA118.004773
Roles of distal aspartate and arginine of B-class dye-decolorizing peroxidase in heterolytic hydrogen peroxide cleavage
Abstract
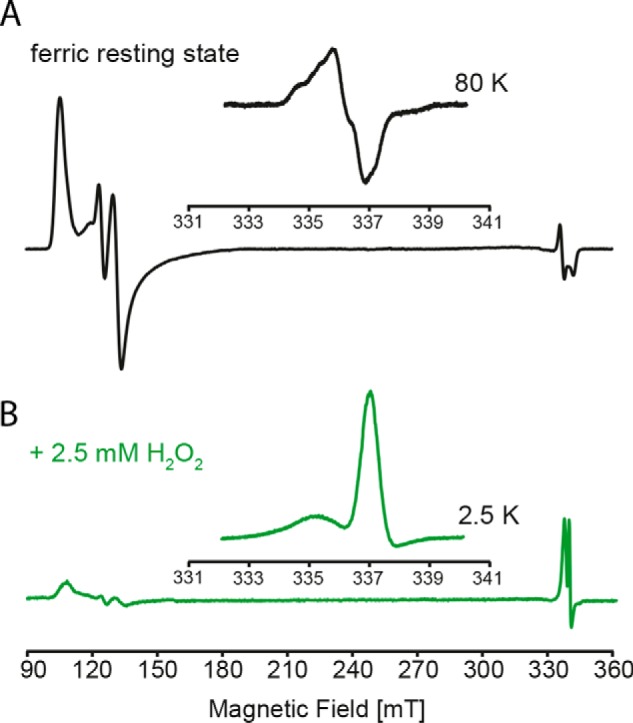
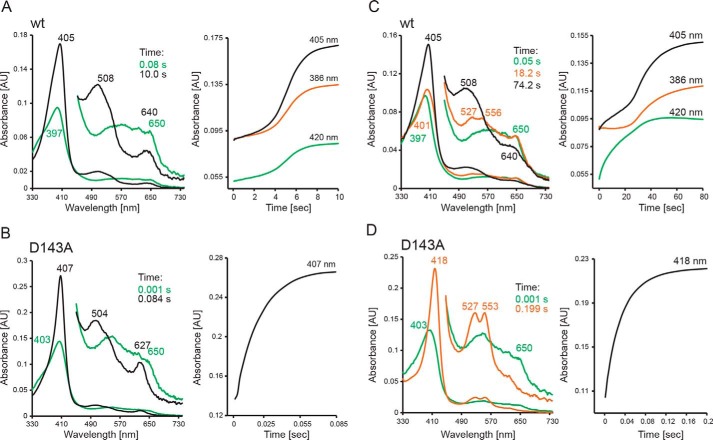
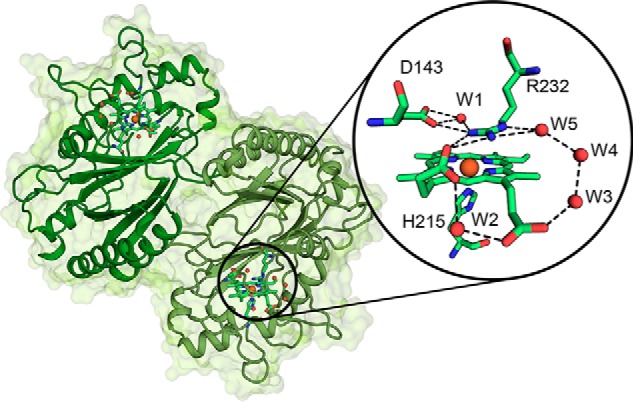
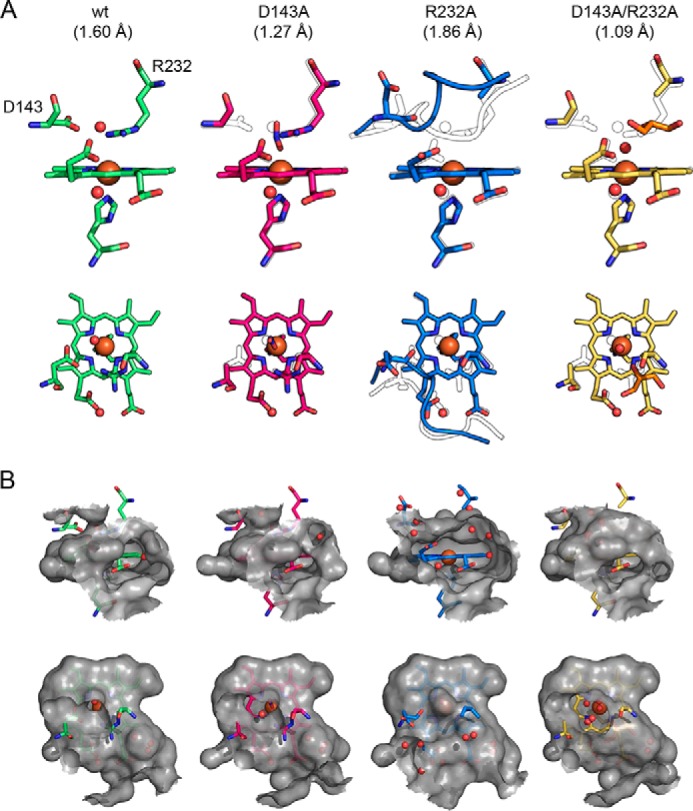
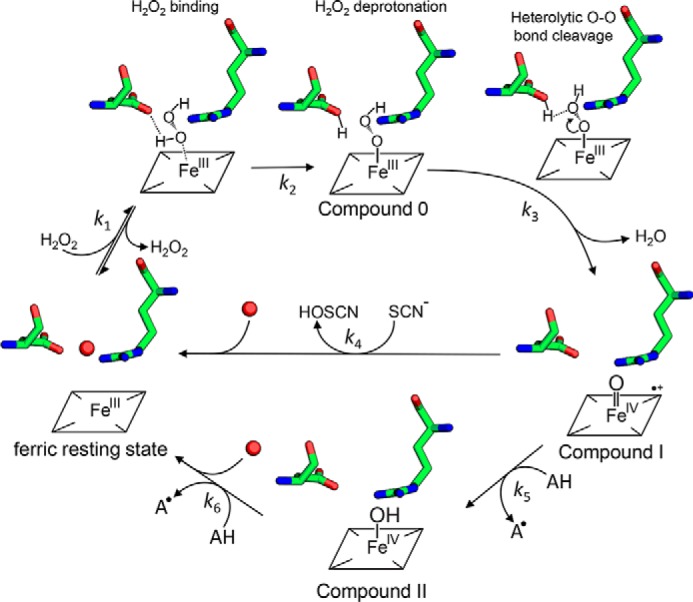
Dye-decolorizing peroxidases (DyPs) represent the most recently classified hydrogen peroxide-dependent heme peroxidase family. Although widely distributed with more than 5000 annotated genes and hailed for their biotechnological potential, detailed biochemical characterization of their reaction mechanism remains limited. Here, we present the high-resolution crystal structures of WT B-class DyP from the pathogenic bacterium Klebsiella pneumoniae (KpDyP) (1.6 Å) and the variants D143A (1.3 Å), R232A (1.9 Å), and D143A/R232A (1.1 Å). We demonstrate the impact of elimination of the DyP-typical, distal residues Asp-143 and Arg-232 on (i) the spectral and redox properties, (ii) the kinetics of heterolytic cleavage of hydrogen peroxide, (iii) the formation of the low-spin cyanide complex, and (iv) the stability and reactivity of an oxoiron(IV)porphyrin π-cation radical (Compound I). Structural and functional studies reveal that the distal aspartate is responsible for deprotonation of H2O2 and for the poor oxidation capacity of Compound I. Elimination of the distal arginine promotes a collapse of the distal heme cavity, including blocking of one access channel and a conformational change of the catalytic aspartate. We also provide evidence of formation of an oxoiron(IV)-type Compound II in KpDyP with absorbance maxima at 418, 527, and 553 nm. In summary, a reaction mechanism of the peroxidase cycle of B-class DyPs is proposed. Our observations challenge the idea that peroxidase activity toward conventional aromatic substrates is related to the physiological roles of B-class DyPs.
Keywords: Compound I; Compound II; Klebsiella pneumonia; X-ray crystallography; electron paramagnetic resonance (EPR); enzyme kinetics; heme; heme peroxidase; oxoiron; pre-steady-state kinetics; site-directed mutagenesis.
© 2018 Pfanzagl et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
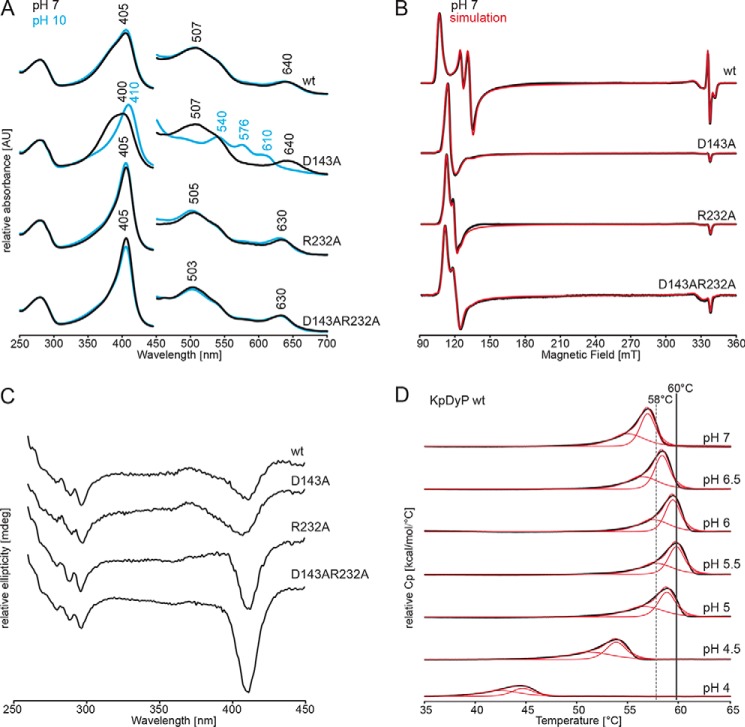
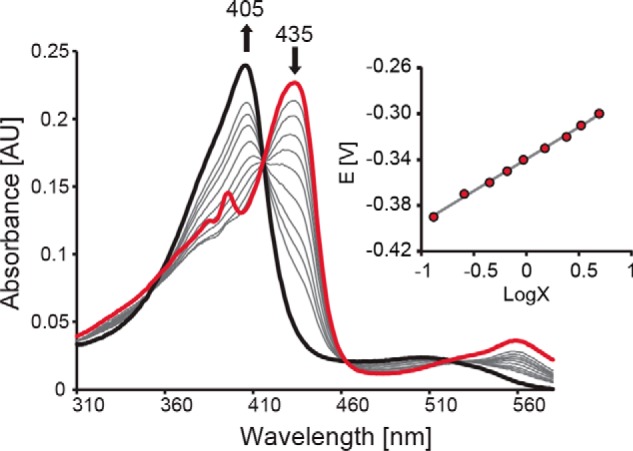
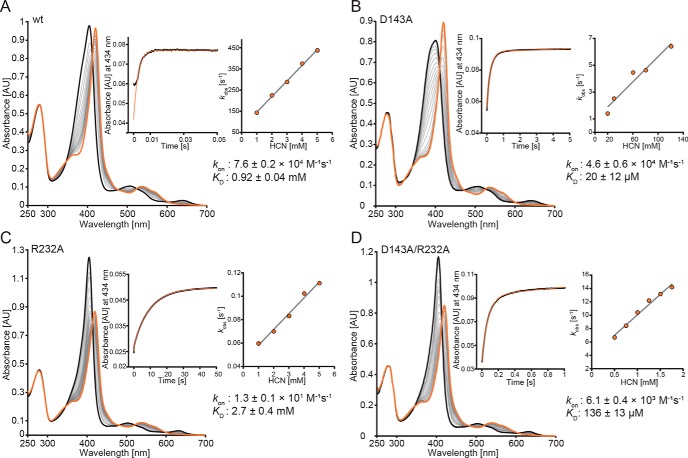
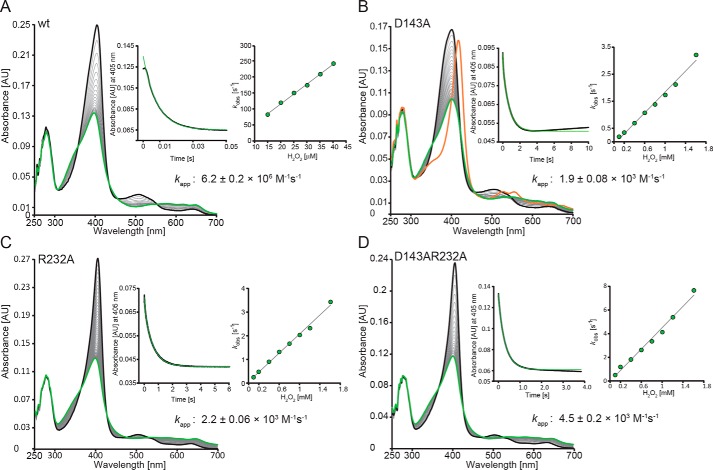
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
