

Emergent simplicity in microbial community assembly
- PMID: 30072533
- PMCID: PMC6405290
- DOI: 10.1126/science.aat1168
Emergent simplicity in microbial community assembly
Abstract
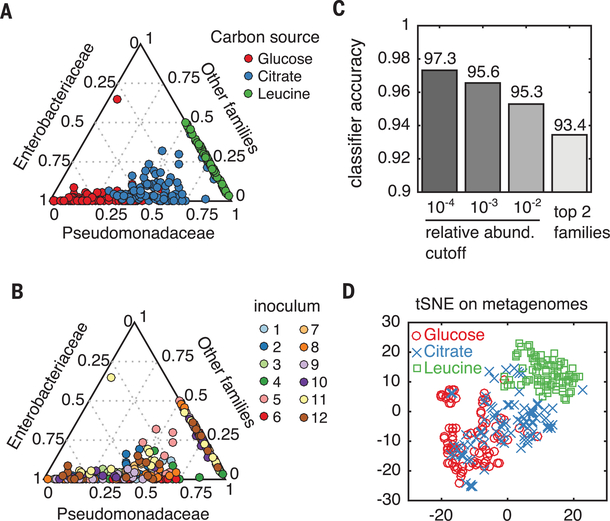
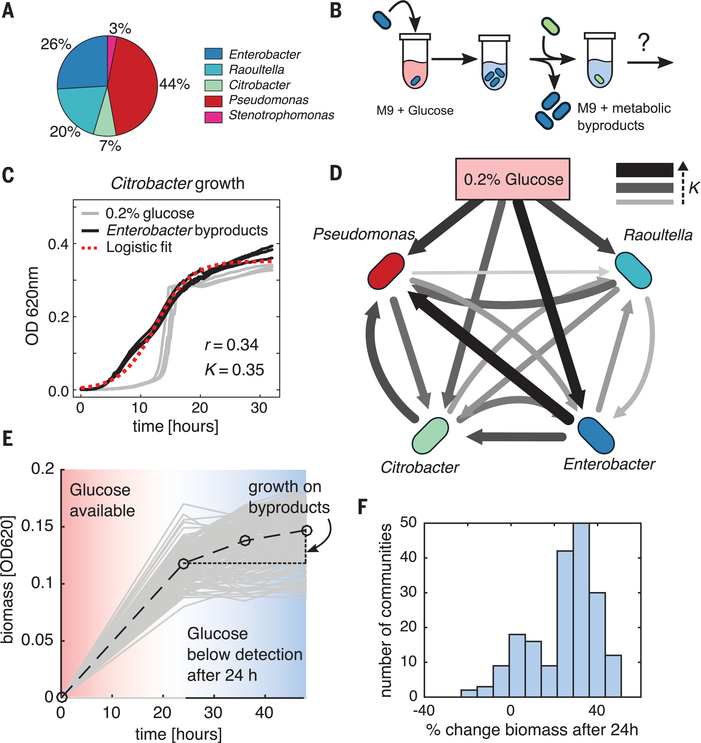
A major unresolved question in microbiome research is whether the complex taxonomic architectures observed in surveys of natural communities can be explained and predicted by fundamental, quantitative principles. Bridging theory and experiment is hampered by the multiplicity of ecological processes that simultaneously affect community assembly in natural ecosystems. We addressed this challenge by monitoring the assembly of hundreds of soil- and plant-derived microbiomes in well-controlled minimal synthetic media. Both the community-level function and the coarse-grained taxonomy of the resulting communities are highly predictable and governed by nutrient availability, despite substantial species variability. By generalizing classical ecological models to include widespread nonspecific cross-feeding, we show that these features are all emergent properties of the assembly of large microbial communities, explaining their ubiquity in natural microbiomes.
Copyright © 2018, American Association for the Advancement of Science.
Conflict of interest statement
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
