

Modeling Tissue and Blood Gas Kinetics in Coastal and Offshore Common Bottlenose Dolphins, Tursiops truncatus
- PMID: 30072907
- PMCID: PMC6060447
- DOI: 10.3389/fphys.2018.00838
Modeling Tissue and Blood Gas Kinetics in Coastal and Offshore Common Bottlenose Dolphins, Tursiops truncatus
Abstract
Bottlenose dolphins (Tursiops truncatus) are highly versatile breath-holding predators that have adapted to a wide range of foraging niches from rivers and coastal ecosystems to deep-water oceanic habitats. Considerable research has been done to understand how bottlenose dolphins manage O2 during diving, but little information exists on other gases or how pressure affects gas exchange. Here we used a dynamic multi-compartment gas exchange model to estimate blood and tissue O2, CO2, and N2 from high-resolution dive records of two different common bottlenose dolphin ecotypes inhabiting shallow (Sarasota Bay) and deep (Bermuda) habitats. The objective was to compare potential physiological strategies used by the two populations to manage shallow and deep diving life styles. We informed the model using species-specific parameters for blood hematocrit, resting metabolic rate, and lung compliance. The model suggested that the known O2 stores were sufficient for Sarasota Bay dolphins to remain within the calculated aerobic dive limit (cADL), but insufficient for Bermuda dolphins that regularly exceeded their cADL. By adjusting the model to reflect the body composition of deep diving Bermuda dolphins, with elevated muscle mass, muscle myoglobin concentration and blood volume, the cADL increased beyond the longest dive duration, thus reflecting the necessary physiological and morphological changes to maintain their deep-diving life-style. The results indicate that cardiac output had to remain elevated during surface intervals for both ecotypes, and suggests that cardiac output has to remain elevated during shallow dives in-between deep dives to allow sufficient restoration of O2 stores for Bermuda dolphins. Our integrated modeling approach contradicts predictions from simple models, emphasizing the complex nature of physiological interactions between circulation, lung compression, and gas exchange.
Keywords: blood gases; decompression sickness; diving physiology; gas exchange; hypoxia; marine mammals; modeling and simulations.
Figures
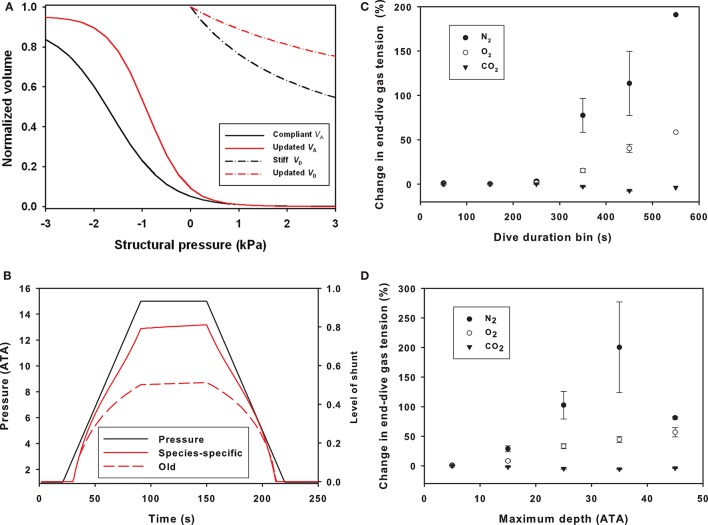
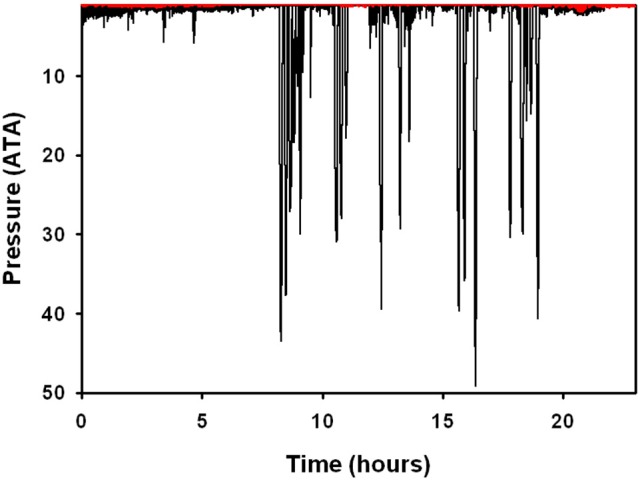
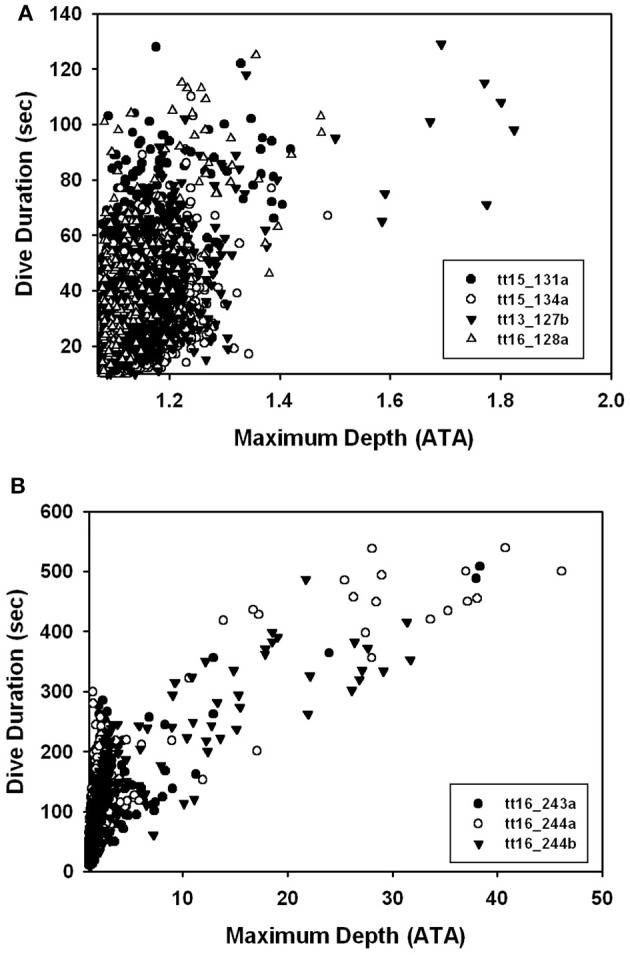
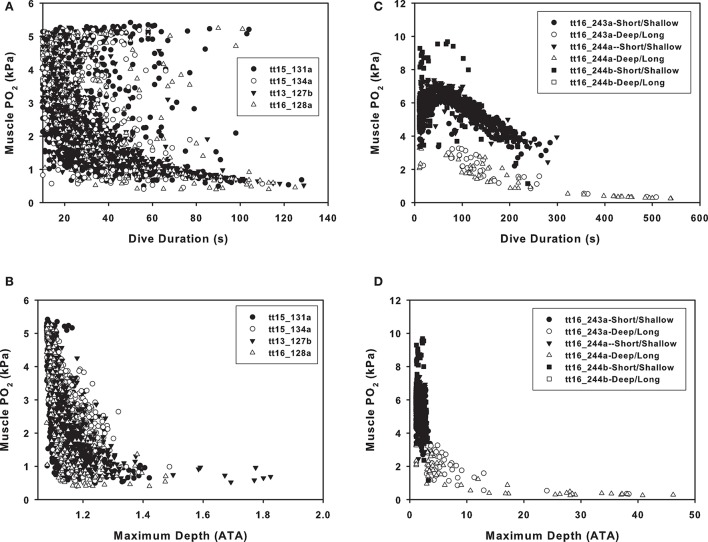
References
-
- Andrews R. D., Jones D. R., Williams J. D., Thorson P. H., Oliver G. W., Costa D. P., et al. . (1997). Heart rates of northern Elephant seals diving at sea and resting on the beach. J. Exp. Biol. 200, 2083–2095. - PubMed
-
- Barros N. B., Wells R. S. (1998). Prey and feeding patterns of resident bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. J. Mam. 79, 1045–1059. 10.2307/1383114 - DOI
-
- Boyd I. L., Woakes A. J., Butler P. J., Davis R. W., Williams T. M. (1995). Validation of heart rate and doubly labelled water as measures of metabolic rate during swimming in California sea lions. Func. Ecol. 9, 151–160. 10.2307/2390559 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources