

The effect of Jun dimerization on neurite outgrowth and motif binding
- PMID: 30077771
- PMCID: PMC6547139
- DOI: 10.1016/j.mcn.2018.08.001
The effect of Jun dimerization on neurite outgrowth and motif binding
Abstract
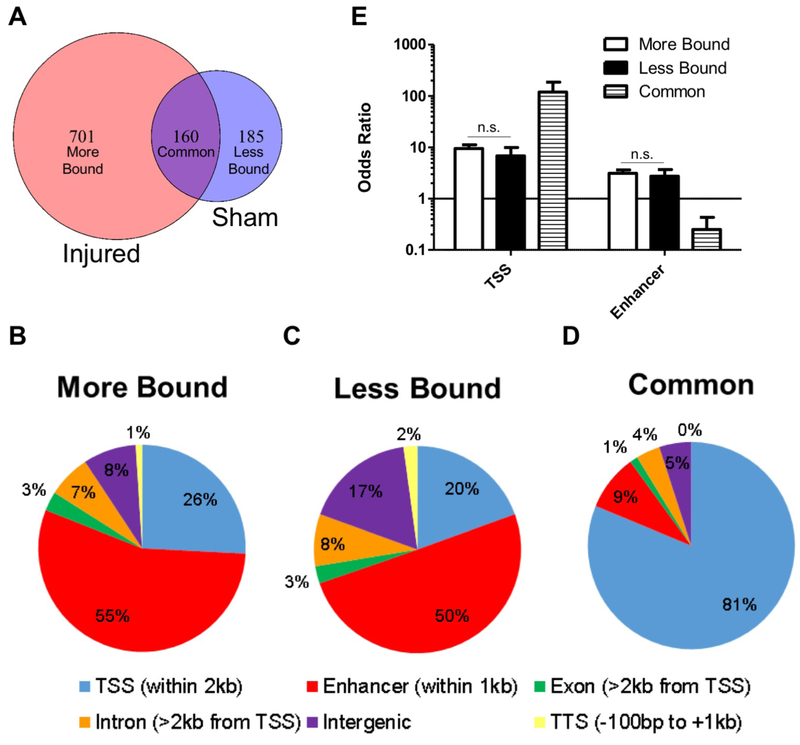
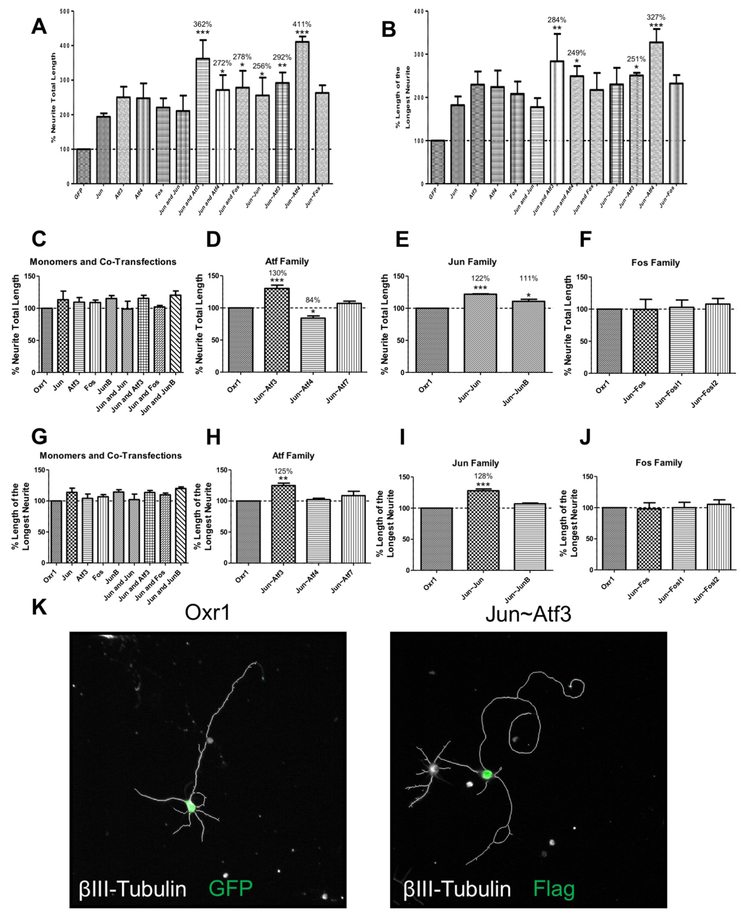
Axon regeneration is a necessary step toward functional recovery after spinal cord injury. The AP-1 transcription factor c-Jun has long been known to play an important role in directing the transcriptional response of Dorsal Root Ganglion (DRG) neurons to peripheral axotomy that results in successful axon regeneration. Here we performed ChIPseq for Jun in mouse DRG neurons after a sciatic nerve crush or sham surgery in order to measure the changes in Jun's DNA binding in response to peripheral axotomy. We found that the majority of Jun's injury-responsive changes in DNA binding occur at putative enhancer elements, rather than proximal to transcription start sites. We also used a series of single polypeptide chain tandem transcription factors to test the effects of different Jun-containing dimers on neurite outgrowth in DRG, cortical and hippocampal neurons. These experiments demonstrated that dimers composed of Jun and Atf3 promoted neurite outgrowth in rat CNS neurons as well as mouse DRG neurons. Our work provides new insight into the mechanisms underlying Jun's role in axon regeneration.
Keywords: AP-1; Chromatin immunoprecipitation; Dorsal Root Ganglion; Enhancer.
Copyright © 2018. Published by Elsevier Inc.
Conflict of interest statement
Competing interests
Conflicts of interest: none
Figures


References
-
- Angel P, Karin M, 1991. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim Biophys Acta. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
