

Resurrecting the Bacterial Tyrosyl-tRNA Synthetase/tRNA Pair for Expanding the Genetic Code of Both E. coli and Eukaryotes
- PMID: 30078635
- PMCID: PMC6456809
- DOI: 10.1016/j.chembiol.2018.07.002
Resurrecting the Bacterial Tyrosyl-tRNA Synthetase/tRNA Pair for Expanding the Genetic Code of Both E. coli and Eukaryotes
Abstract
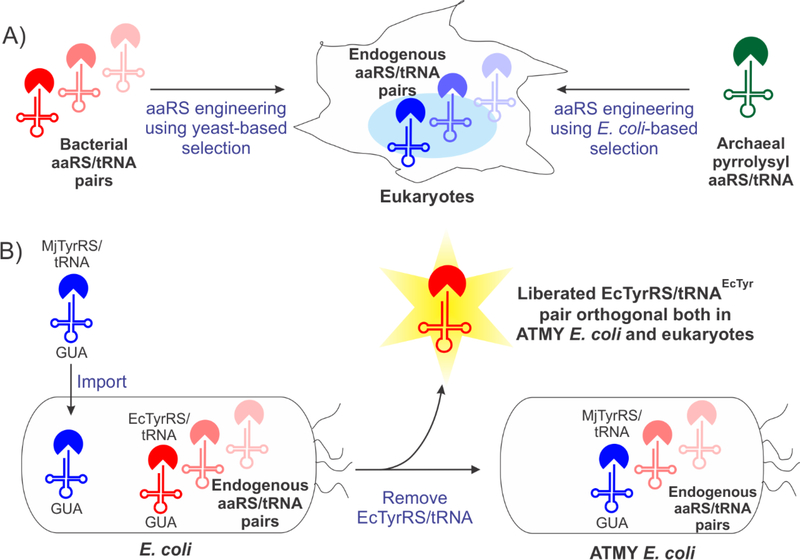
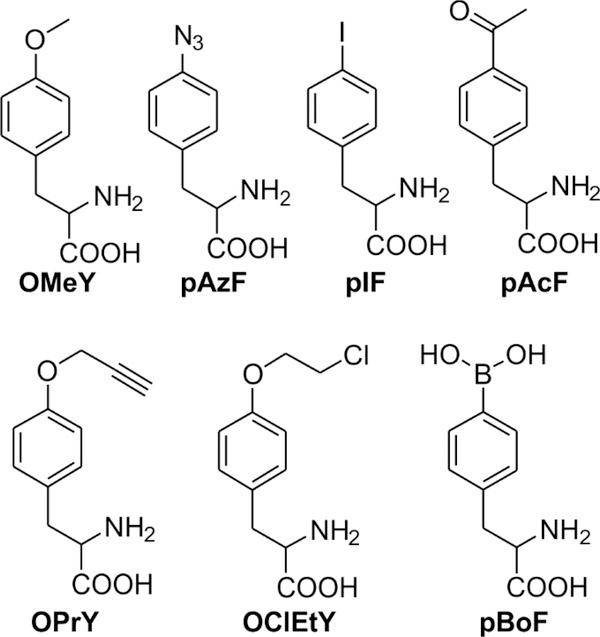
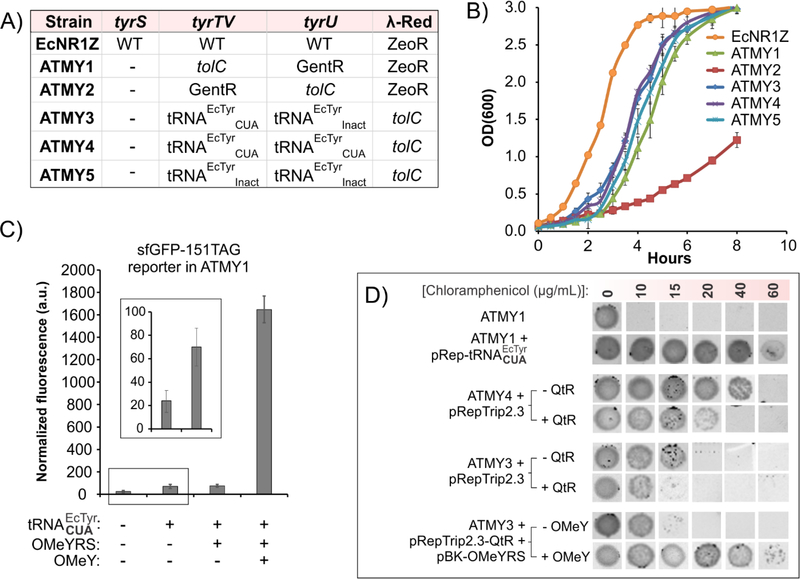
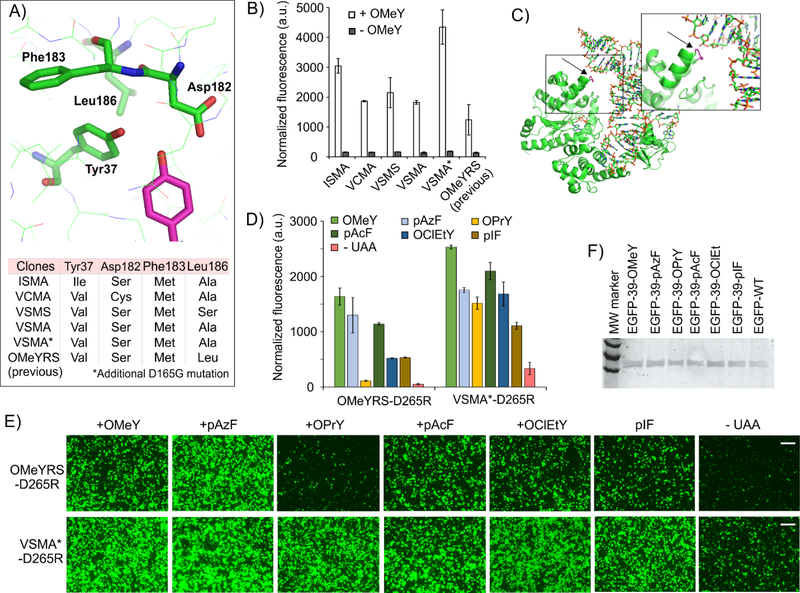
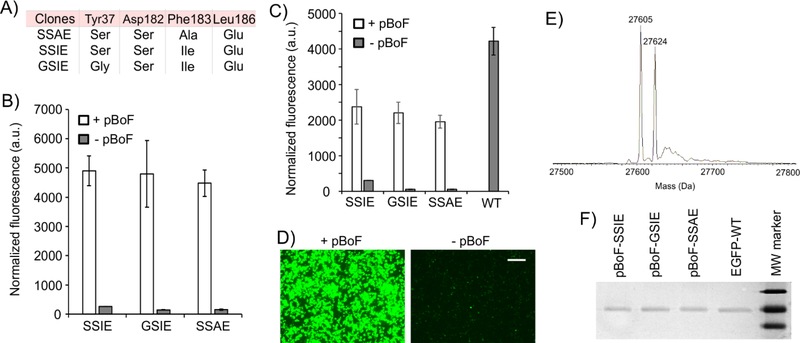
The bacteria-derived tyrosyl-tRNA synthetase (TyrRS)/tRNA pair was first used for unnatural amino acid (Uaa) mutagenesis in eukaryotic cells over 15 years ago. It provides an ideal platform to genetically encode numerous useful Uaas in eukaryotes. However, this pair has been engineered to charge only a small collection of Uaas to date. Development of Uaa-selective variants of this pair has been limited by technical challenges associated with a yeast-based directed evolution platform, which is currently required to alter its substrate specificity. Here we overcome this limitation by enabling its directed evolution in an engineered strain of E. coli (ATMY), where the endogenous TyrRS/tRNA pair has been functionally replaced with an archaeal counterpart. The facile E. coli-based selection system enabled rapid engineering of this pair to develop variants that selectively incorporate various Uaas, including p-boronophenylalanine, into proteins expressed in mammalian cells as well as in the ATMY strain of E. coli.
Keywords: engineered bacterial strains; eukaryotic genetic code expansion; genetic code expansion; mammalian genetic code expansion; non-canonical amino acids; unnatural amino acids.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
