

Dual Roles of Poly(dA:dT) Tracts in Replication Initiation and Fork Collapse
- PMID: 30078706
- PMCID: PMC6591735
- DOI: 10.1016/j.cell.2018.07.011
Dual Roles of Poly(dA:dT) Tracts in Replication Initiation and Fork Collapse
Abstract
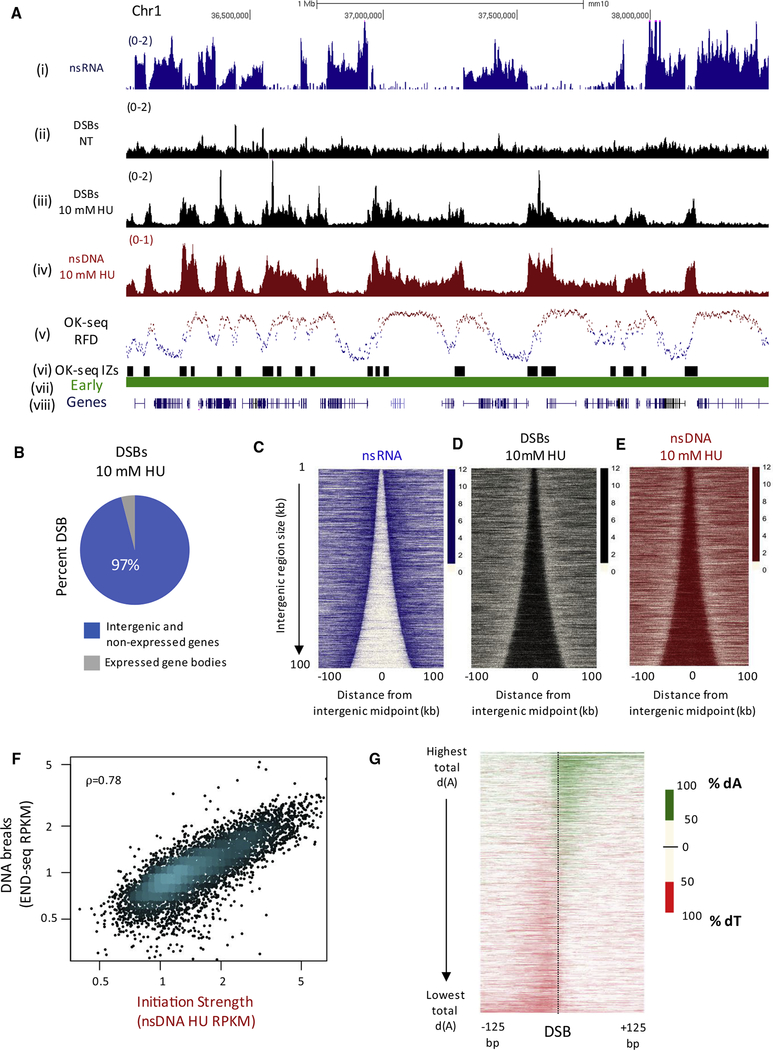
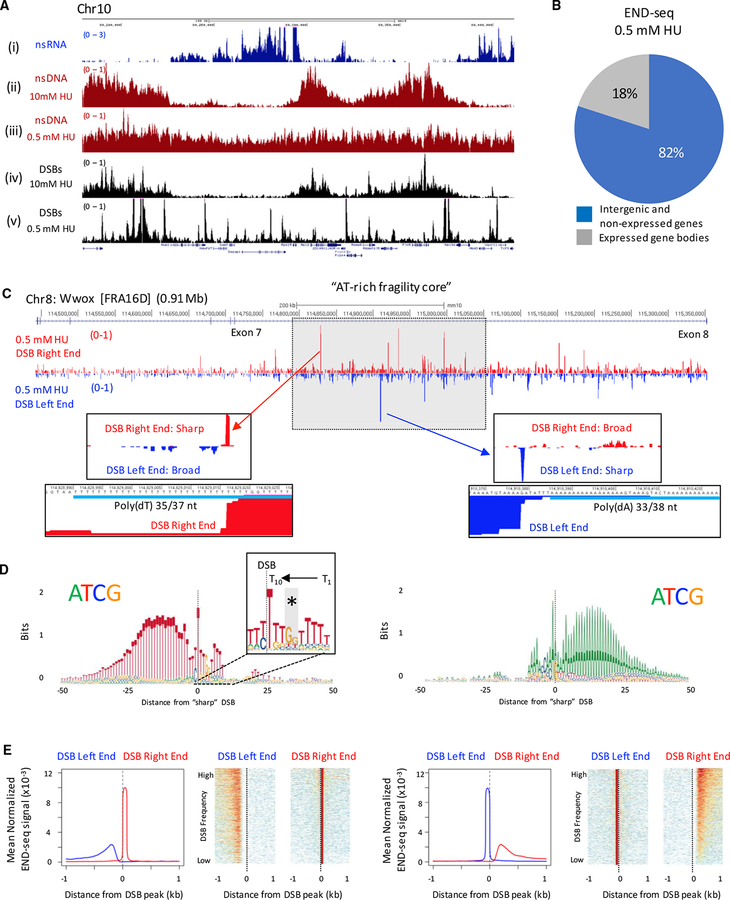
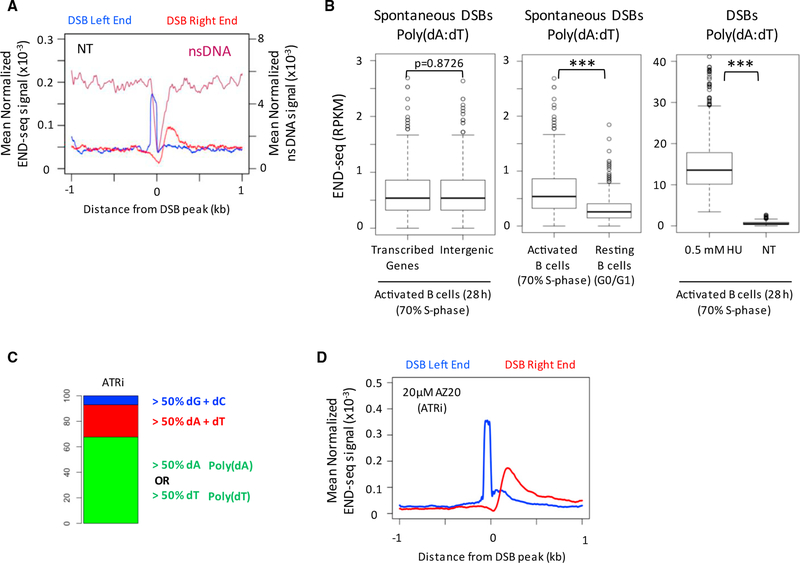
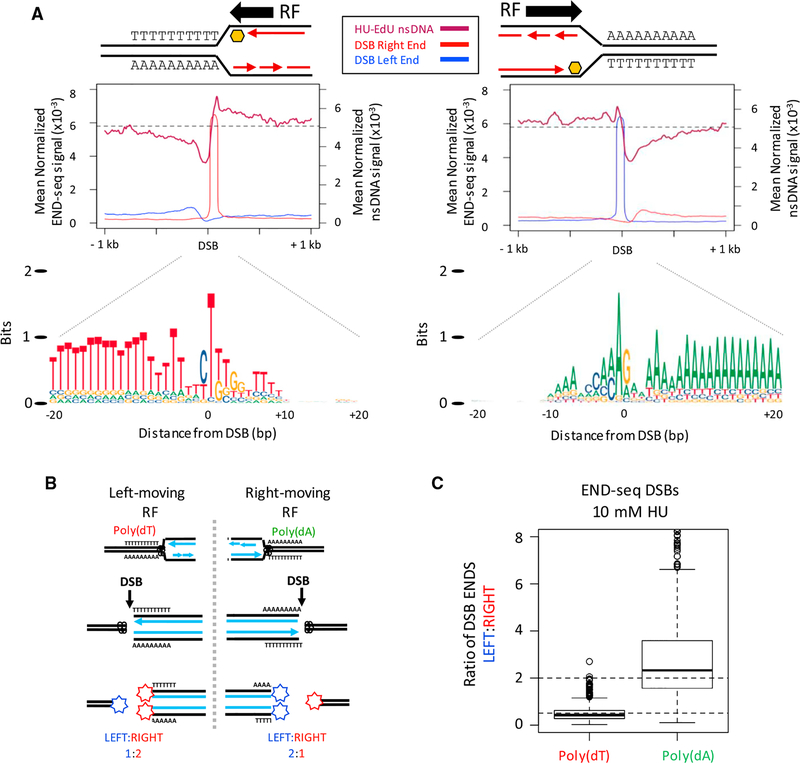
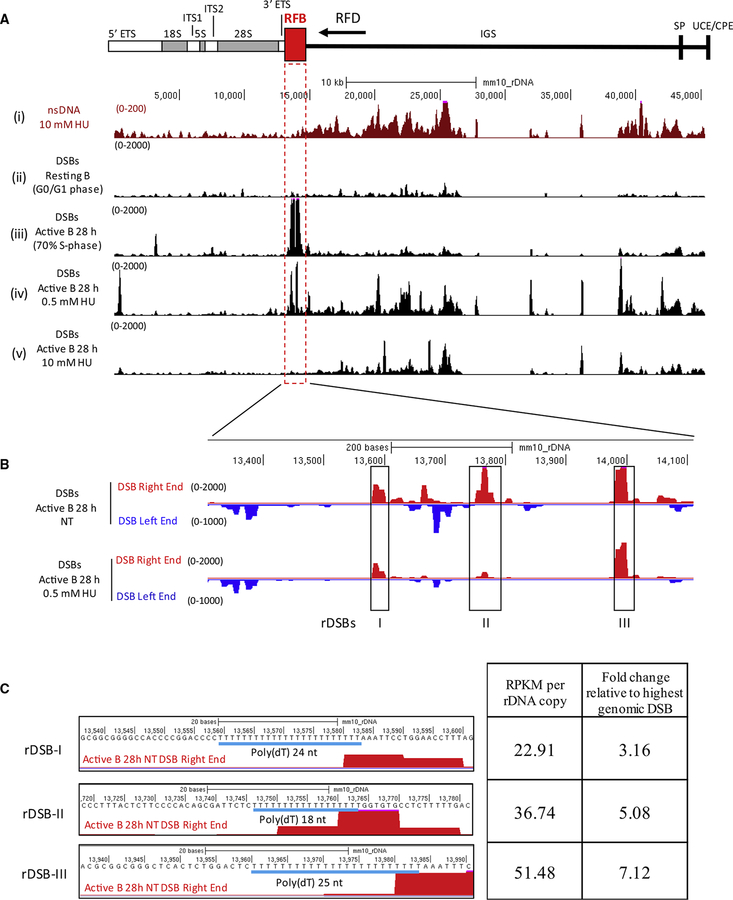
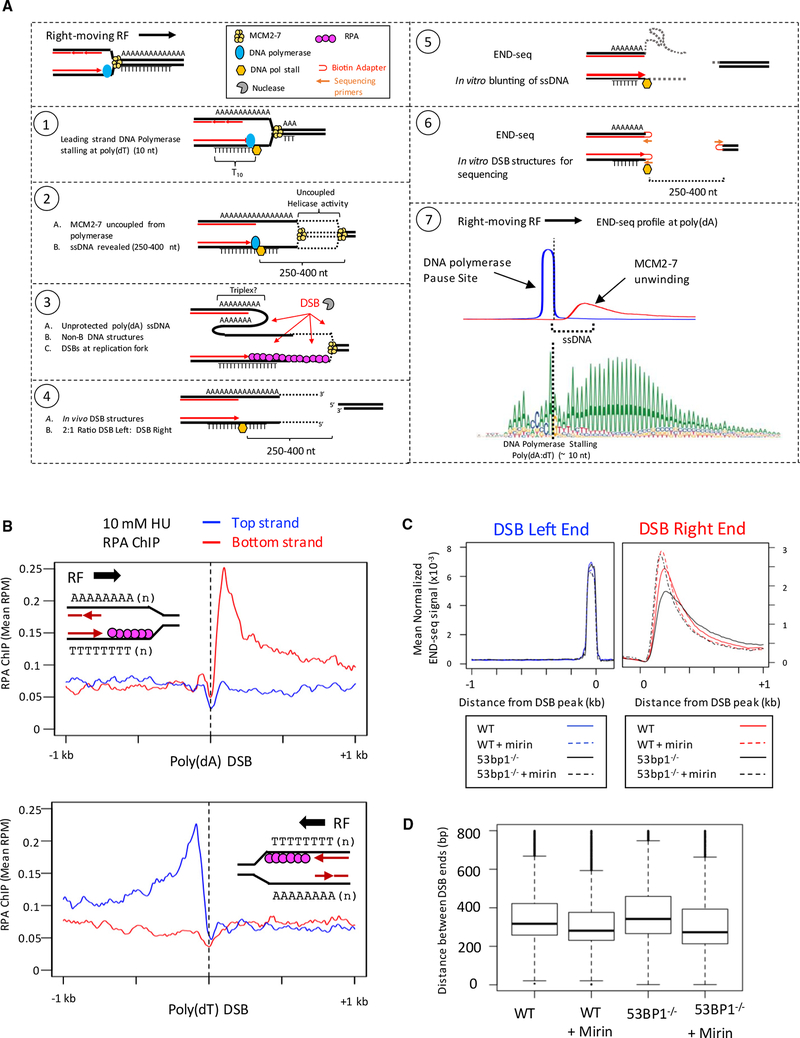
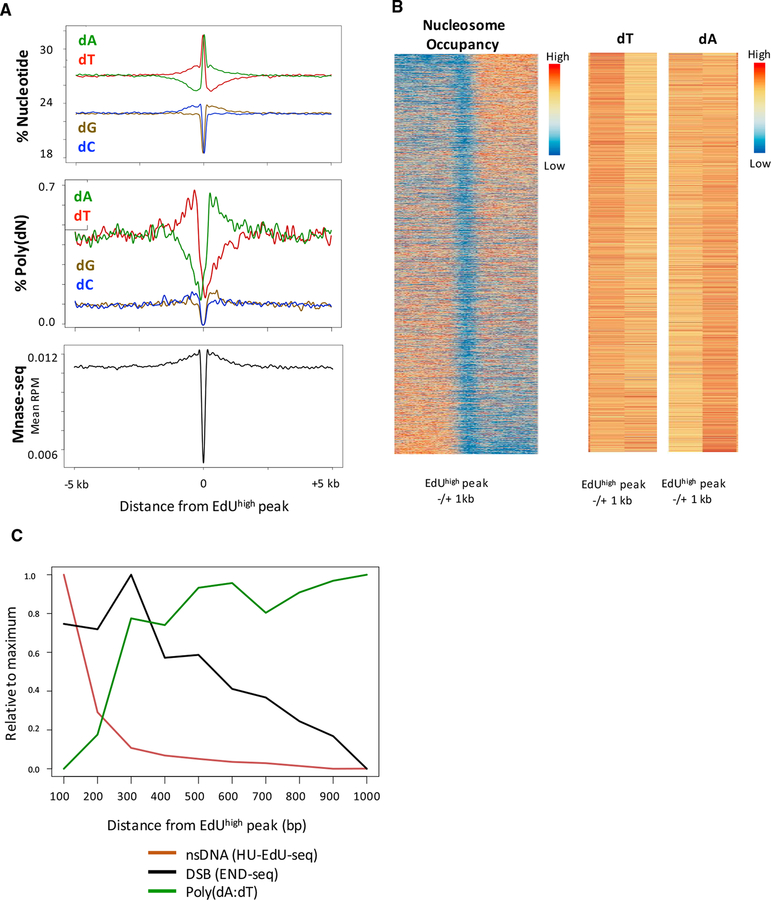
Replication origins, fragile sites, and rDNA have been implicated as sources of chromosomal instability. However, the defining genomic features of replication origins and fragile sites are among the least understood elements of eukaryote genomes. Here, we map sites of replication initiation and breakage in primary cells at high resolution. We find that replication initiates between transcribed genes within nucleosome-depleted structures established by long asymmetrical poly(dA:dT) tracts flanking the initiation site. Paradoxically, long (>20 bp) (dA:dT) tracts are also preferential sites of polar replication fork stalling and collapse within early-replicating fragile sites (ERFSs) and late-replicating common fragile sites (CFSs) and at the rDNA replication fork barrier. Poly(dA:dT) sequences are fragile because long single-strand poly(dA) stretches at the replication fork are unprotected by the replication protein A (RPA). We propose that the evolutionary expansion of poly(dA:dT) tracts in eukaryotic genomes promotes replication initiation, but at the cost of chromosome fragility.
Keywords: DNA breaks; fragile sites; genome instability; poly(dA:dT) tracts; replication fork barrier; replication origins; replication stress; ribosomal DNA.
Published by Elsevier Inc.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Comment in
-
Poly(dA:dT) make it and break it.Nat Rev Mol Cell Biol. 2018 Oct;19(10):619. doi: 10.1038/s41580-018-0054-6. Nat Rev Mol Cell Biol. 2018. PMID: 30111873 No abstract available.
-
Getting a Genomic View of DNA Replication Stress.Mol Cell. 2018 Oct 18;72(2):201-203. doi: 10.1016/j.molcel.2018.10.001. Mol Cell. 2018. PMID: 30340018
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
