

Cicero Predicts cis-Regulatory DNA Interactions from Single-Cell Chromatin Accessibility Data
- PMID: 30078726
- PMCID: PMC6582963
- DOI: 10.1016/j.molcel.2018.06.044
Cicero Predicts cis-Regulatory DNA Interactions from Single-Cell Chromatin Accessibility Data
Abstract
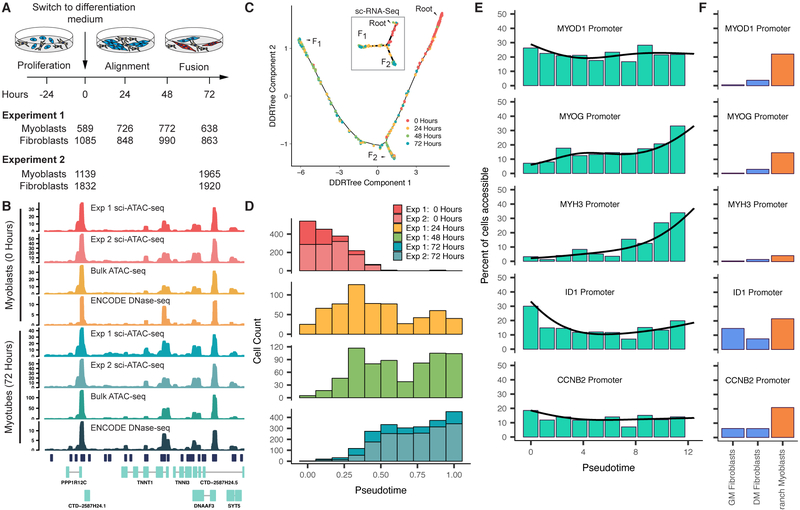
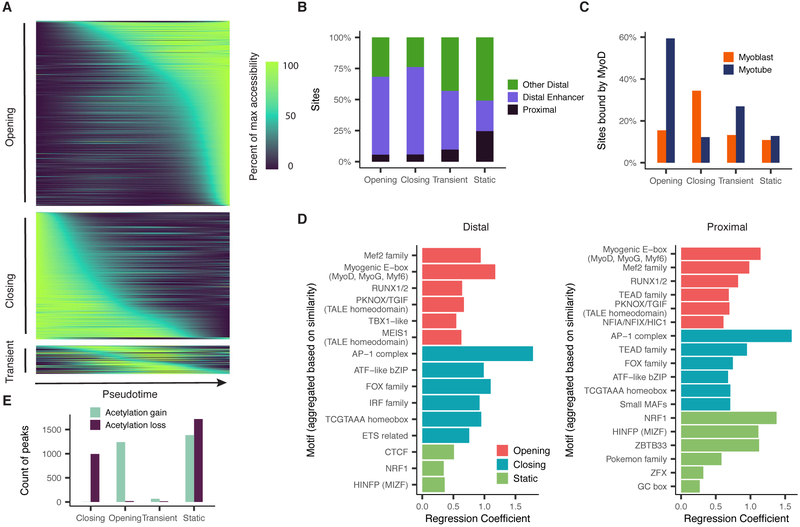
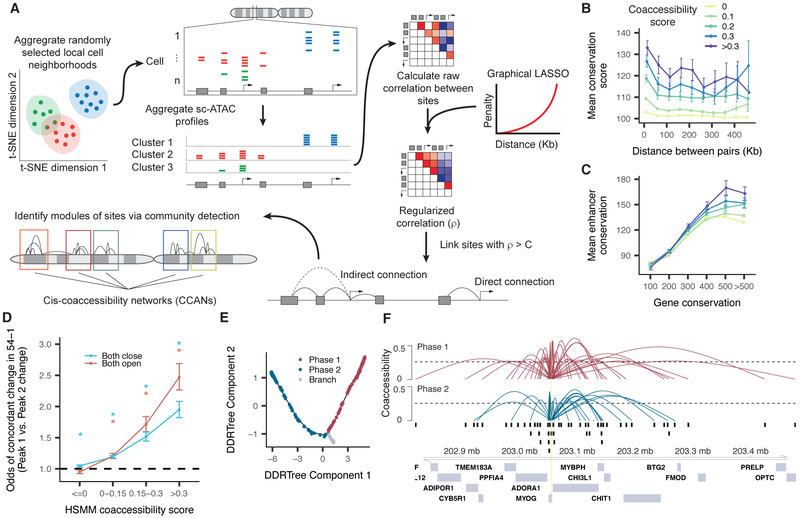
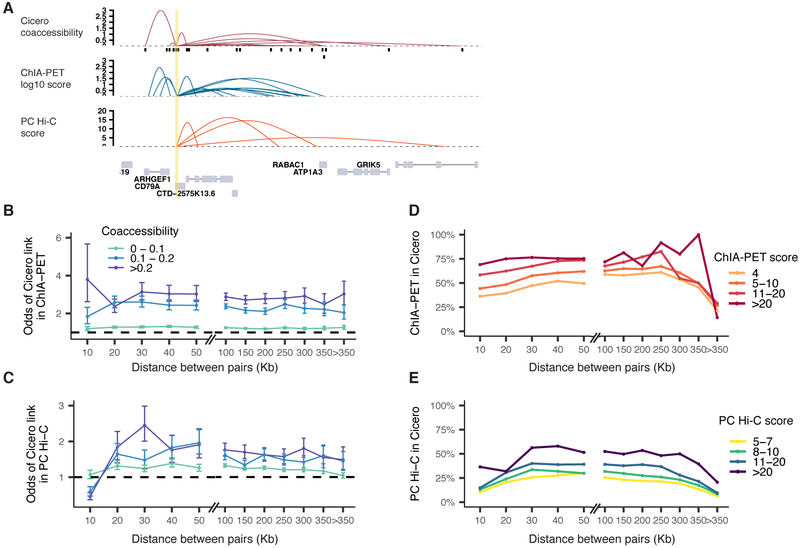
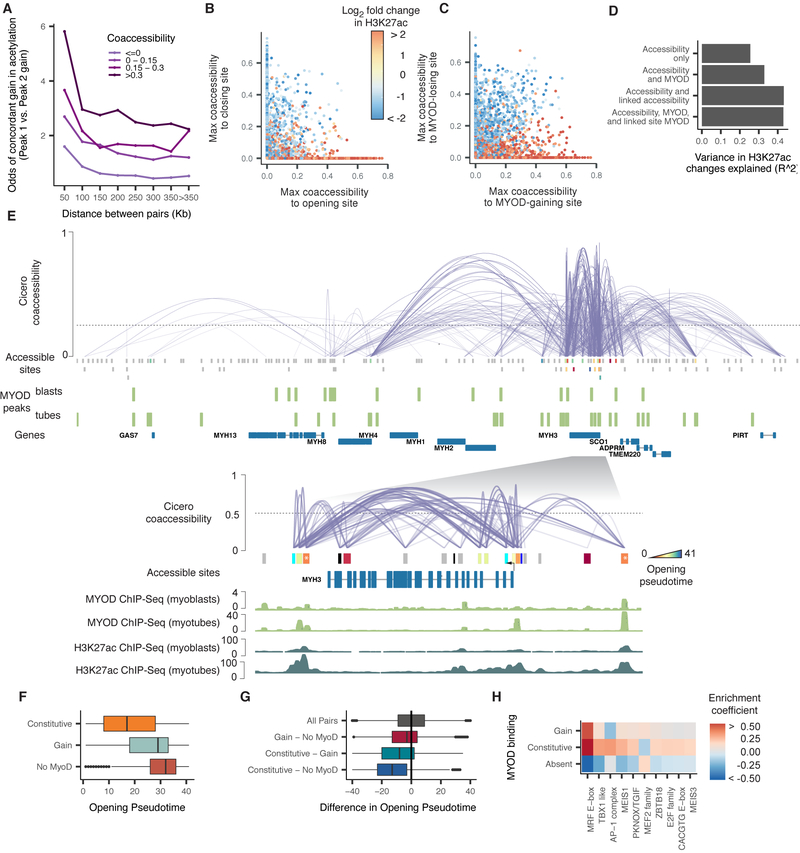
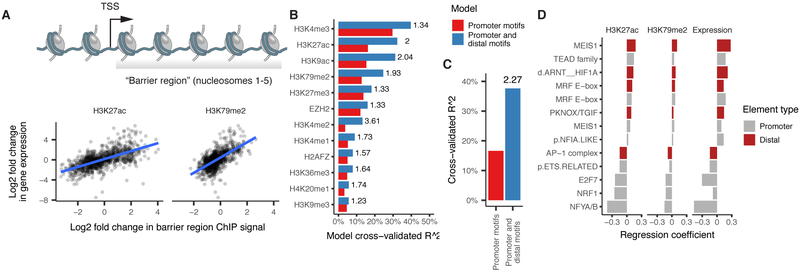
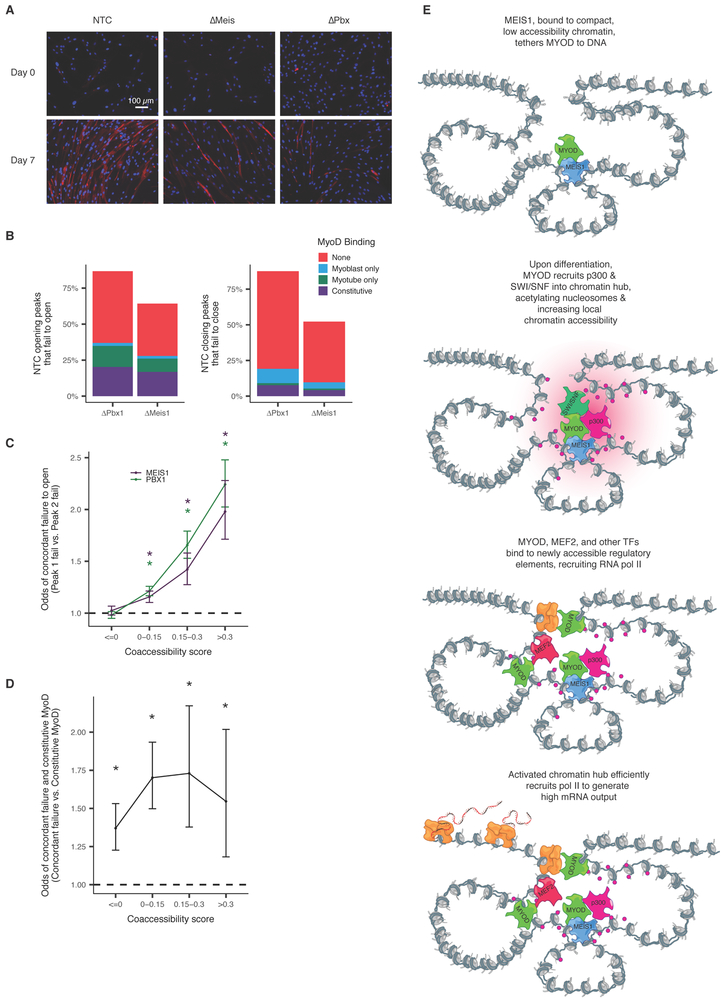
Linking regulatory DNA elements to their target genes, which may be located hundreds of kilobases away, remains challenging. Here, we introduce Cicero, an algorithm that identifies co-accessible pairs of DNA elements using single-cell chromatin accessibility data and so connects regulatory elements to their putative target genes. We apply Cicero to investigate how dynamically accessible elements orchestrate gene regulation in differentiating myoblasts. Groups of Cicero-linked regulatory elements meet criteria of "chromatin hubs"-they are enriched for physical proximity, interact with a common set of transcription factors, and undergo coordinated changes in histone marks that are predictive of changes in gene expression. Pseudotemporal analysis revealed that most DNA elements remain in chromatin hubs throughout differentiation. A subset of elements bound by MYOD1 in myoblasts exhibit early opening in a PBX1- and MEIS1-dependent manner. Our strategy can be applied to dissect the architecture, sequence determinants, and mechanisms of cis-regulation on a genome-wide scale.
Keywords: ATAC-seq; chromatin accessibility; co-accessibility; gene regulation; machine learning; myoblast differentiation; single-cell.
Copyright © 2018. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
LC and FJS have declare competing financial interests in the form of stock ownership and paid employment by Illumina, Inc. One or more embodiments of one or more patents and patent applications filed by Illumina may encompass the methods, reagents, and the data disclosed in this manuscript.
Figures
References
-
- Benezra R, Davis RL, Lockshon D, Turner DL, and Weintraub H (1990). The protein Id: a negative regulator of helix-loop-helix DNA binding proteins. Cell 61, 49–59. - PubMed
-
- Berkes CA, Bergstrom DA, Penn BH, Seaver KJ, Knoepfler PS, and Tapscott SJ (2004). Pbx marks genes for activation by MyoD indicating a role for a homeodomain protein in establishing myogenic potential. Mol. Cell 14, 465–477. - PubMed
-
- Beygelzimer A, Kakadet S, Langford J, Arya S, Mount D, and Li S (2013). FNN: fast nearest neighbor search algorithms and applications. R Package Version 1.
-
- Blondel VD, Guillaume J-L, Lambiotte R, and Lefebvre E (2008). Fast unfolding of communities in large networks.
-
- Breiman L (1996). Bagging predictors. Mach. Learn. 24, 123–140.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
