

MICU1 imparts the mitochondrial uniporter with the ability to discriminate between Ca2+ and Mn2+
- PMID: 30082385
- PMCID: PMC6112746
- DOI: 10.1073/pnas.1807811115
MICU1 imparts the mitochondrial uniporter with the ability to discriminate between Ca2+ and Mn2+
Abstract
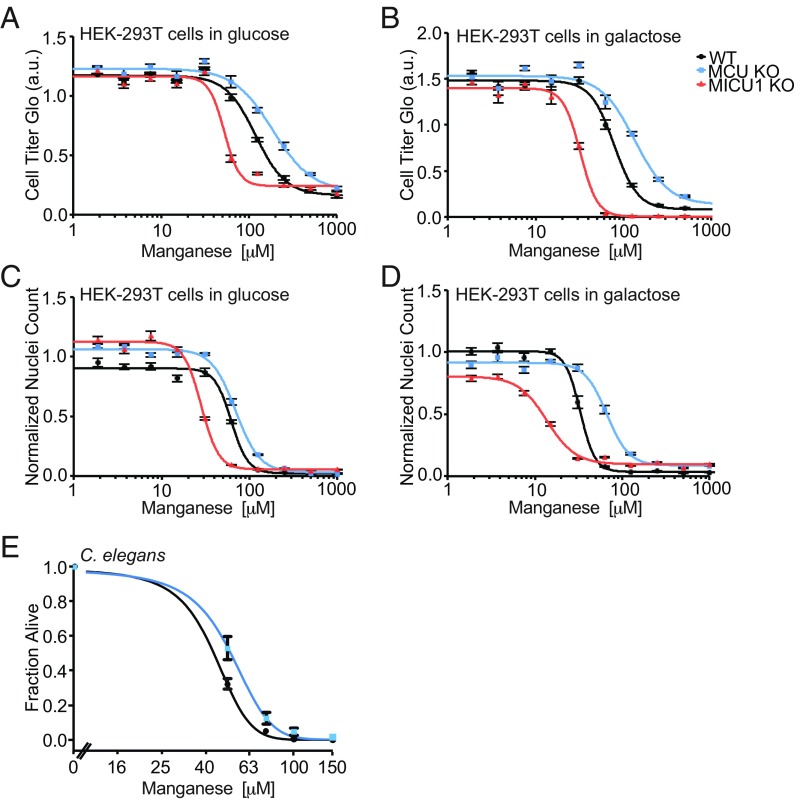
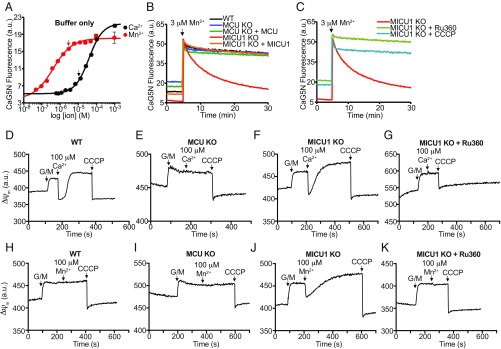
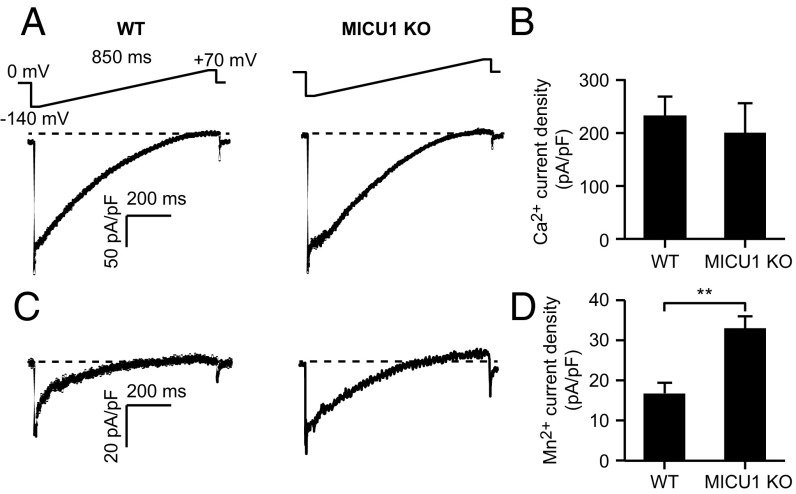
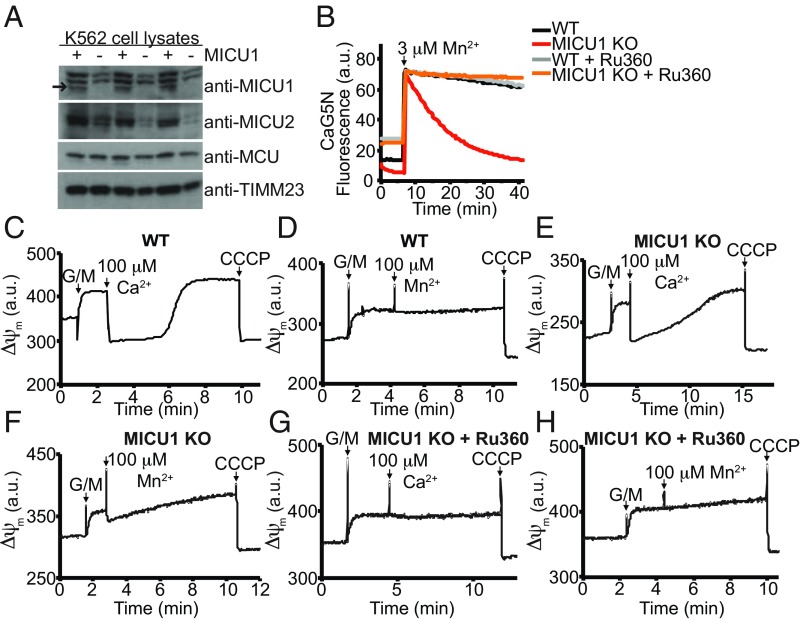
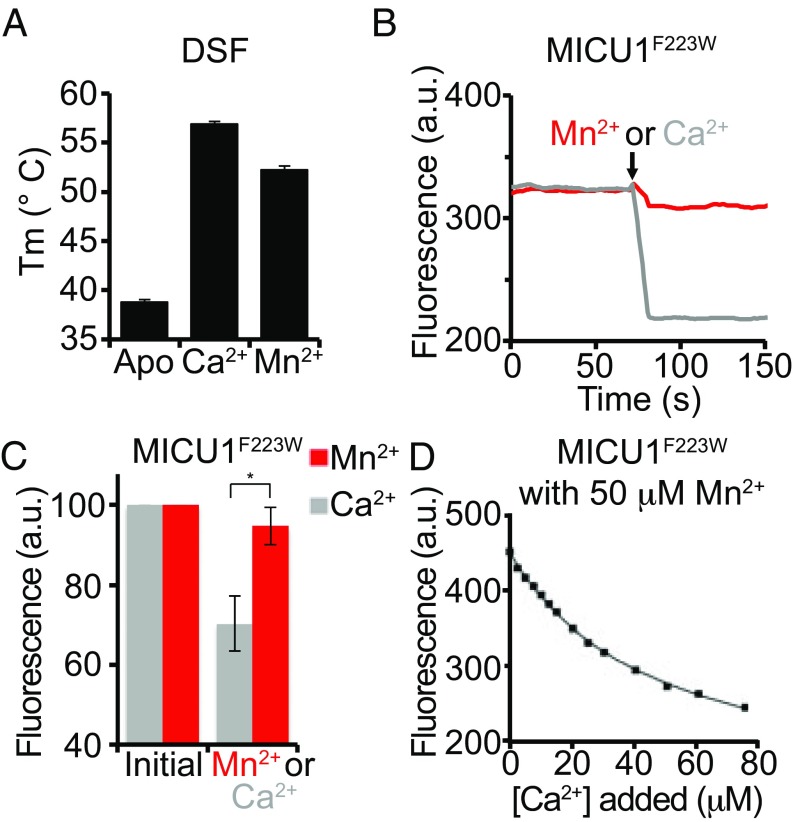
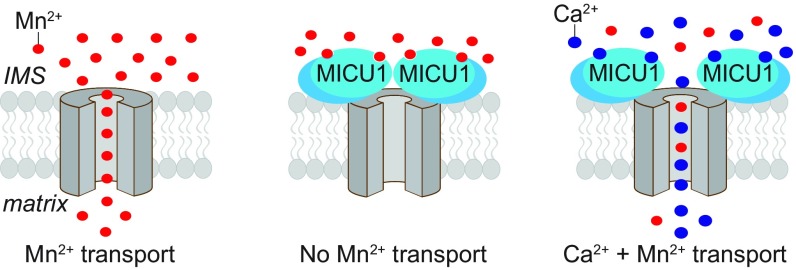
The mitochondrial uniporter is a Ca2+-activated Ca2+ channel complex that displays exceptionally high conductance and selectivity. Here, we report cellular metal toxicity screens highlighting the uniporter's role in Mn2+ toxicity. Cells lacking the pore-forming uniporter subunit, MCU, are more resistant to Mn2+ toxicity, while cells lacking the Ca2+-sensing inhibitory subunit, MICU1, are more sensitive than the wild type. Consistent with these findings, Caenorhabditis elegans lacking the uniporter's pore have increased resistance to Mn2+ toxicity. The chemical-genetic interaction between uniporter machinery and Mn2+ toxicity prompted us to hypothesize that Mn2+ can indeed be transported by the uniporter's pore, but this transport is prevented by MICU1. To this end, we demonstrate that, in the absence of MICU1, both Mn2+ and Ca2+ can pass through the uniporter, as evidenced by mitochondrial Mn2+ uptake assays, mitochondrial membrane potential measurements, and mitoplast electrophysiology. We show that Mn2+ does not elicit the conformational change in MICU1 that is physiologically elicited by Ca2+, preventing Mn2+ from inducing the pore opening. Our work showcases a mechanism by which a channel's auxiliary subunit can contribute to its apparent selectivity and, furthermore, may have implications for understanding how manganese contributes to neurodegenerative disease.
Keywords: EF hand; calcium; manganese; neurodegeneration; selectivity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kirichok Y, Krapivinsky G, Clapham DE. The mitochondrial calcium uniporter is a highly selective ion channel. Nature. 2004;427:360–364. - PubMed
-
- Sather WA, McCleskey EW. Permeation and selectivity in calcium channels. Annu Rev Physiol. 2003;65:133–159. - PubMed
-
- Maynard LS, Cotzias GC. The partition of manganese among organs and intracellular organelles of the rat. J Biol Chem. 1955;214:489–495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
