

The taste of ribonucleosides: Novel macronutrients essential for larval growth are sensed by Drosophila gustatory receptor proteins
- PMID: 30086130
- PMCID: PMC6080749
- DOI: 10.1371/journal.pbio.2005570
The taste of ribonucleosides: Novel macronutrients essential for larval growth are sensed by Drosophila gustatory receptor proteins
Abstract
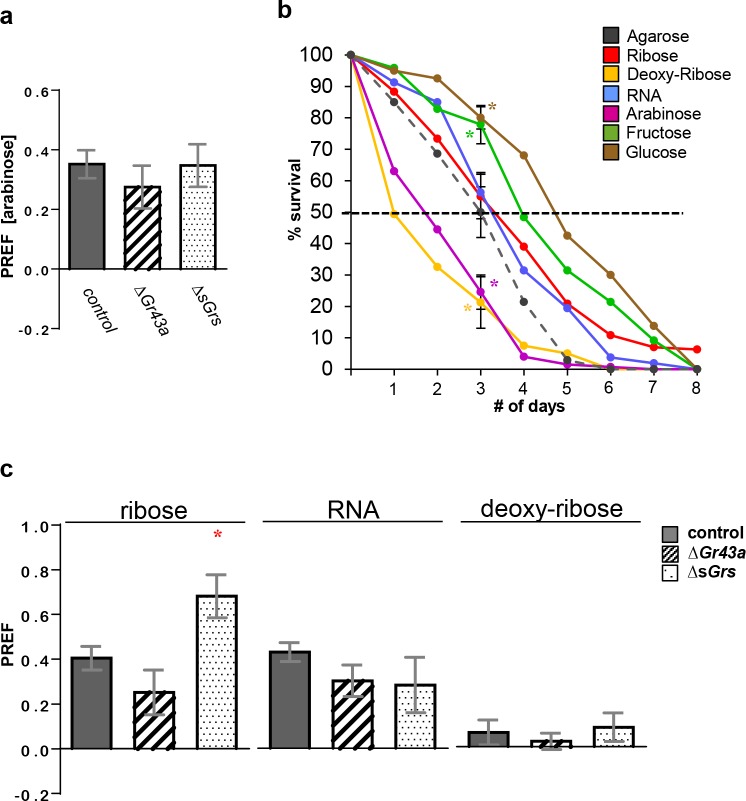
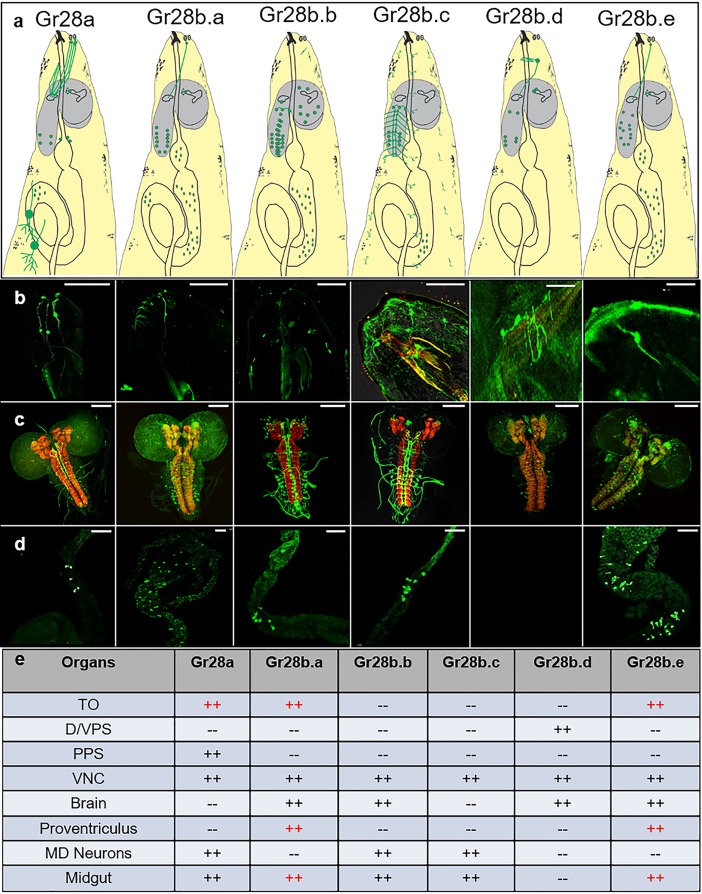
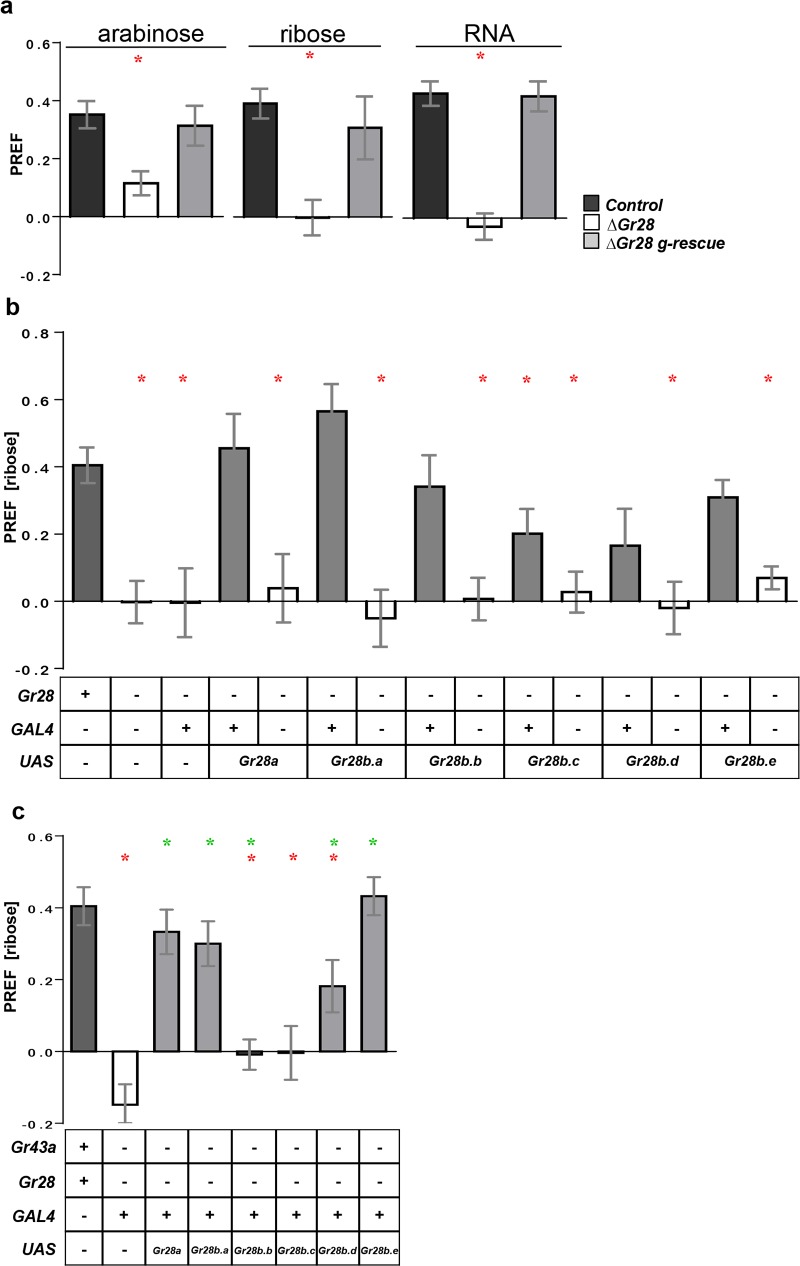
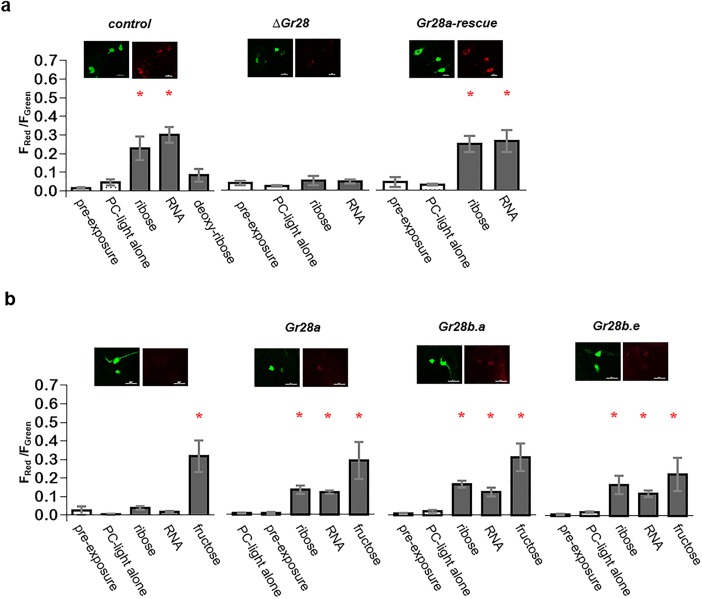
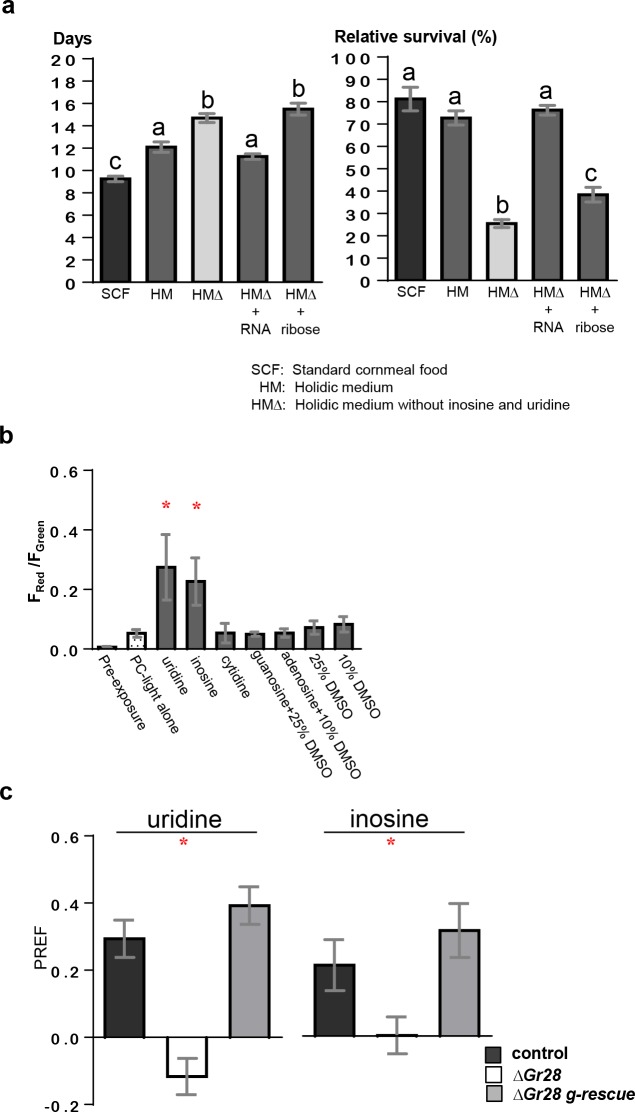
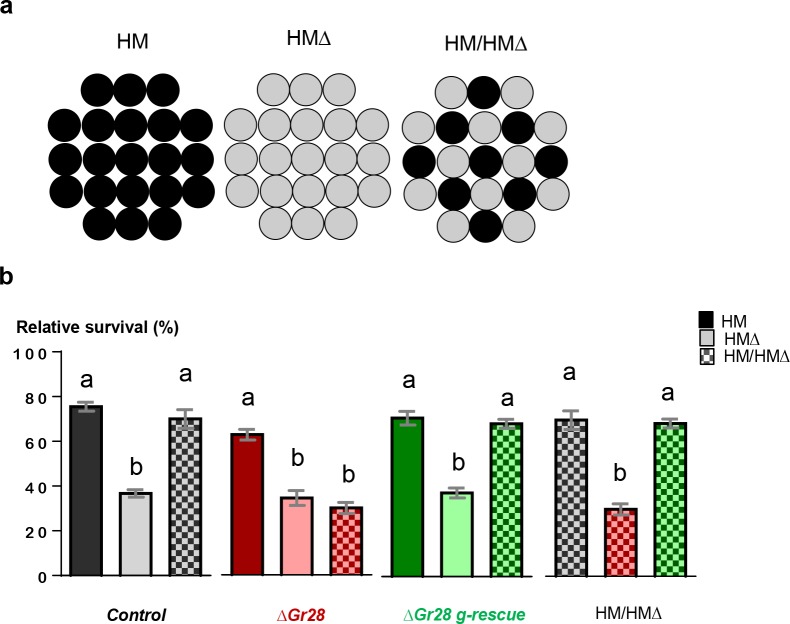
Animals employ various types of taste receptors to identify and discriminate between different nutritious food chemicals. These macronutrients are thought to fall into 3 major groups: carbohydrates/sugars, proteins/amino acids, and fats. Here, we report that Drosophila larvae exhibit a novel appetitive feeding behavior towards ribose, ribonucleosides, and RNA. We identified members of the gustatory receptor (Gr) subfamily 28 (Gr28), expressed in both external and internal chemosensory neurons as molecular receptors necessary for cellular and appetitive behavioral responses to ribonucleosides and RNA. Specifically, behavioral preference assays show that larvae are strongly attracted to ribose- or RNA-containing agarose in a Gr28-dependent manner. Moreover, Ca2+ imaging experiments reveal that Gr28a-expressing taste neurons are activated by ribose, RNA and some ribonucleosides and that these responses can be conveyed to Gr43aGAL4 fructose-sensing neurons by expressing single members of the Gr28 gene family. Lastly, we establish a critical role in behavioral fitness for the Gr28 genes by showing that Gr28 mutant larvae exhibit low survival rates when challenged to find ribonucleosides in food. Together, our work identifies a novel taste modality dedicated to the detection of RNA and ribonucleosides, nutrients that are essential for survival during the accelerated growth phase of Drosophila larvae.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
