

ATP in the tumour microenvironment drives expression of nfP2X7, a key mediator of cancer cell survival
- PMID: 30087439
- PMCID: PMC6328436
- DOI: 10.1038/s41388-018-0426-6
ATP in the tumour microenvironment drives expression of nfP2X7, a key mediator of cancer cell survival
Abstract
The ATP-gated receptor P2X7 is expressed in multiple malignant tumours including neuroblastoma, melanoma, prostate, lung and breast. P2X7 has a significant role in mediating diverse cell responses, which upon dysregulation are associated with tumour initiation and development. The rapid, ATP-mediated activation of P2X7 induces a fast-inward cation current in cells. However, prolonged ATP-mediated activation of P2X7 leads to formation of a pore that increases membrane permeability and eventually causes cell death. This presents a potential paradox, as the tumour microenvironment contains extracellular ATP at levels sufficient to activate the P2X7 pore and trigger cell death. However, P2X7 expression is associated with enhanced cancer cell survival, proliferation and metastatic potential. At least one distinct conformational form of P2X7, termed non-pore functional P2X7 (nfP2X7), has been described, which is not able to form a functional pore. We demonstrate for the first time in this study that exposure to a high ATP concentration, equivalent to those measured in the tumour microenvironment, drives nfP2X7 expression and also that nfP2X7 is essential for tumour cell survival. We show that monoclonal antibodies raised against a P2X7 amino acid sequence (200-216), whose conformation is distinct from that of wild-type (WT) P2X7, bind specifically to nfP2X7 expressed on the surface of tumour cells. We also show that nfP2X7 is broadly expressed in patient-derived tumour sections from a wide range of cancers. Therefore, antibodies raised against E200 provide tools that can differentiate between forms of the P2X7 receptor that have a key role in cancer.
Conflict of interest statement
SMG, CJO, SM and RL are employees of Biosceptre (UK) Limited, a company commercialising nfP2X7-targeted therapies. ALP and PB were contracted by Biosceptre (UK) Limited as part of Charles River Discovery Research Services Germany GmbH (formerly named Oncotest GmbH). SH is BBSRC CASE PhD student partly funded by Biosceptre (UK) Limited. FDV serves as a member of Biosceptre International Limited’s Scientific Advisory Board. SF has no conflict of interest.
Figures
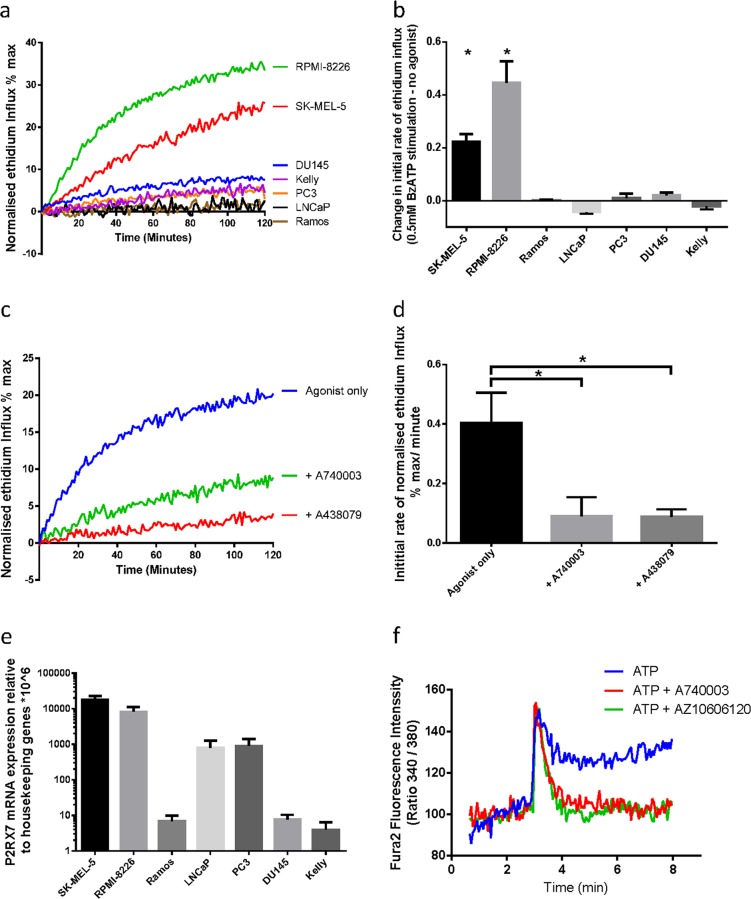
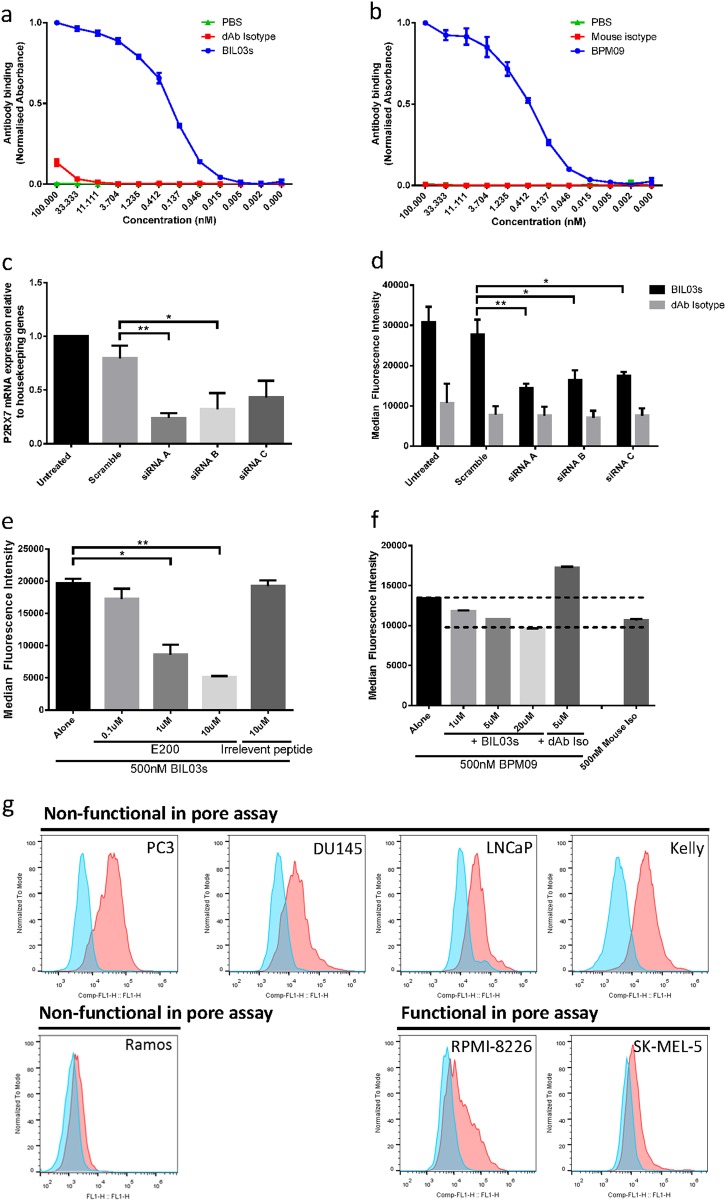
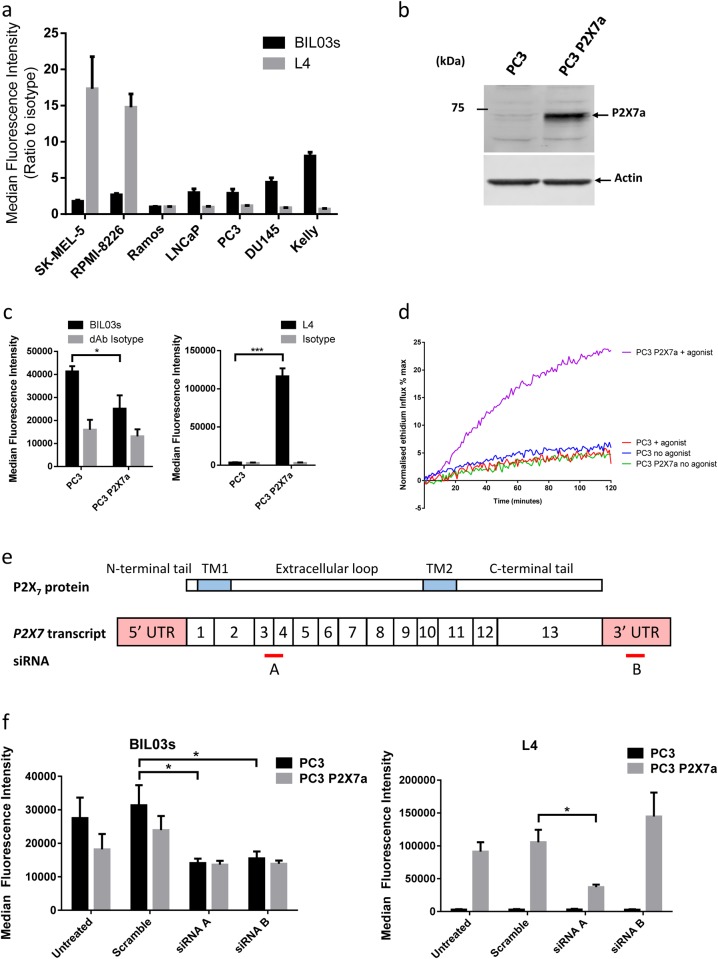
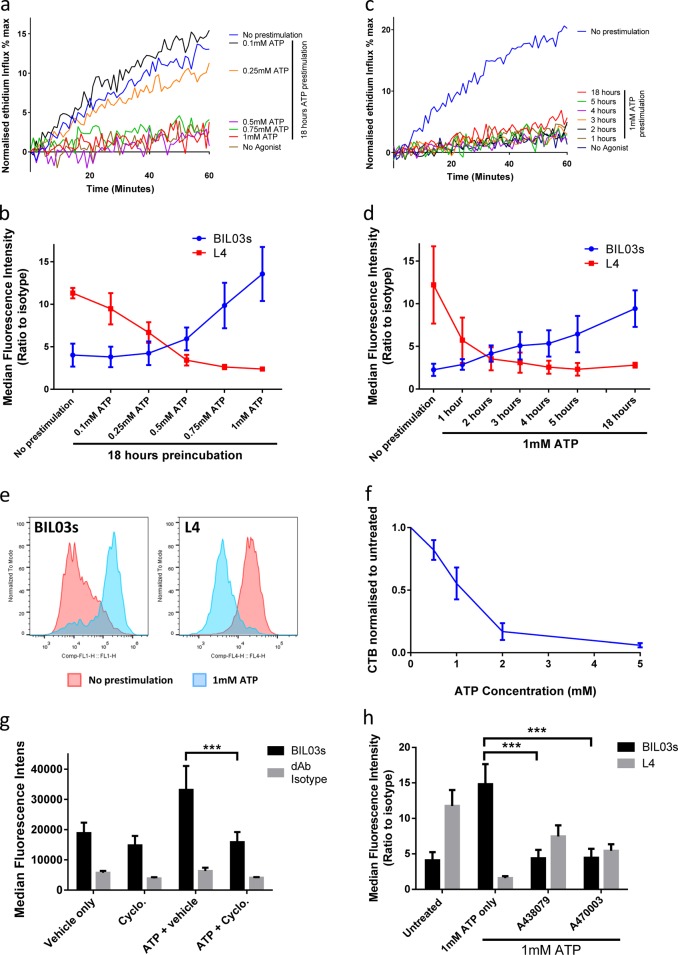
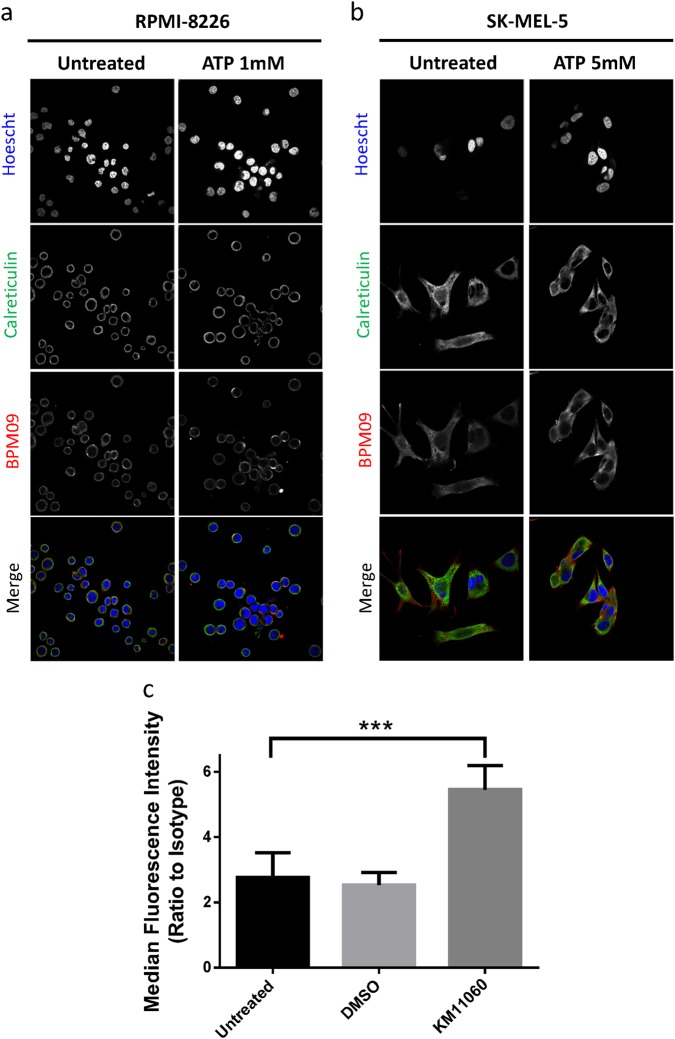
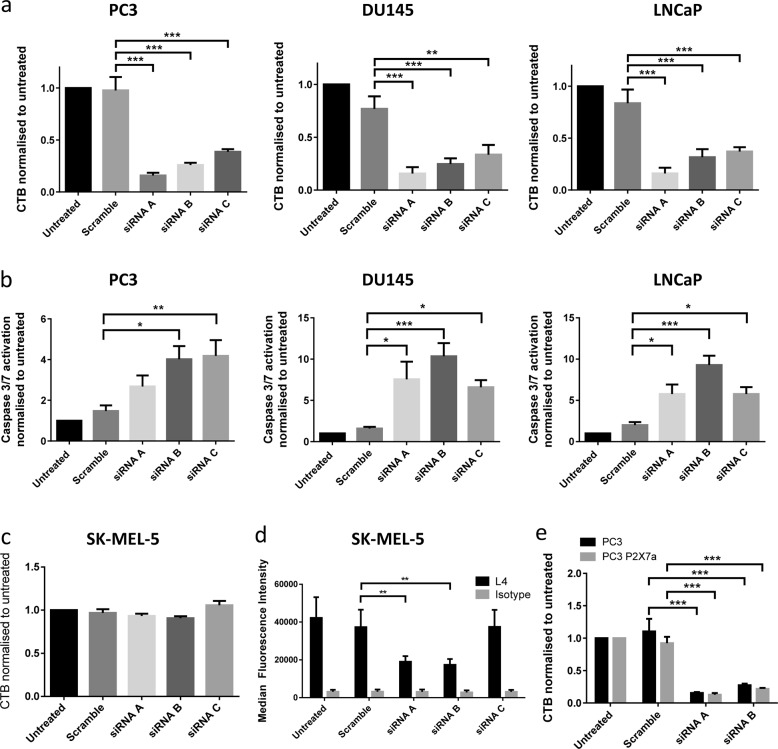
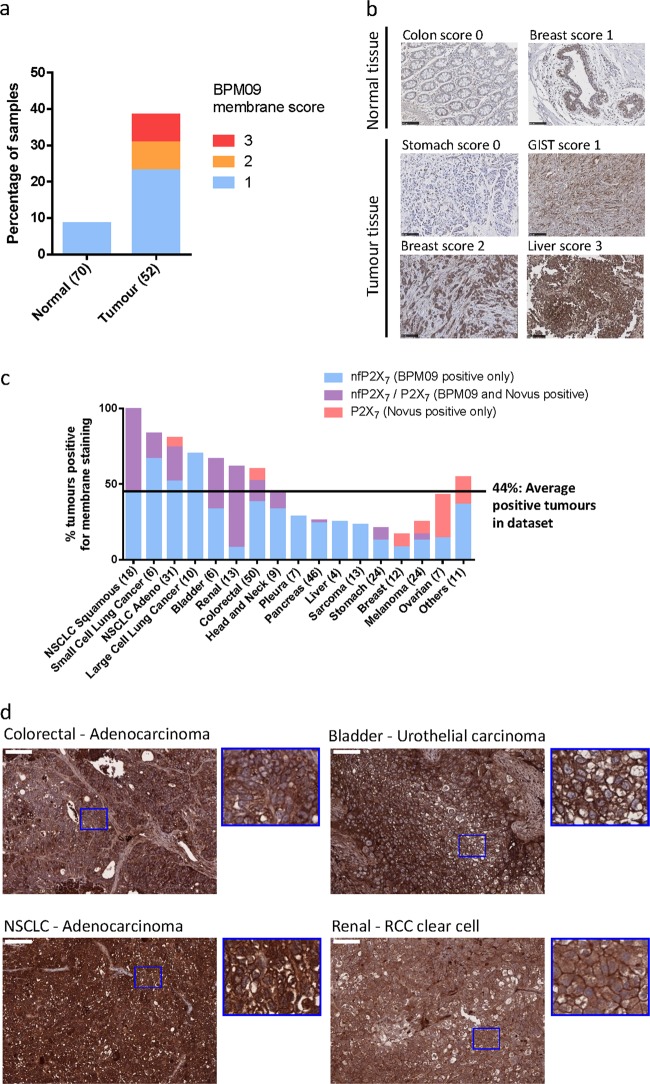
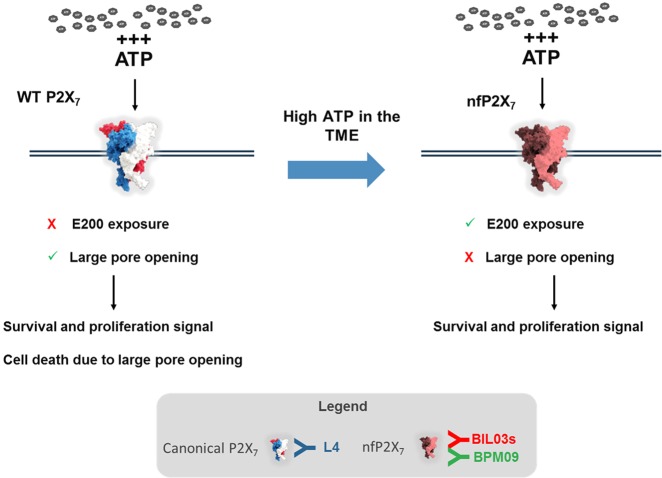
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
