

Heterochromatin and RNAi regulate centromeres by protecting CENP-A from ubiquitin-mediated degradation
- PMID: 30089114
- PMCID: PMC6101405
- DOI: 10.1371/journal.pgen.1007572
Heterochromatin and RNAi regulate centromeres by protecting CENP-A from ubiquitin-mediated degradation
Abstract
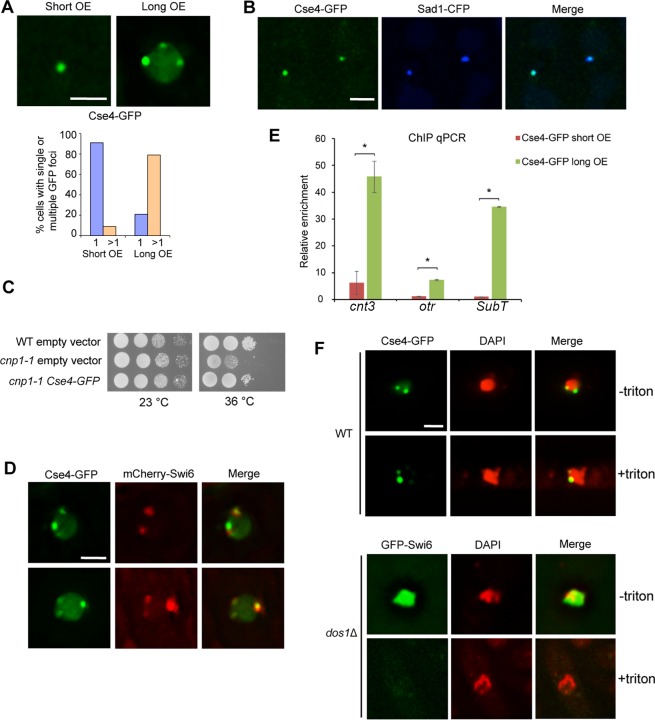
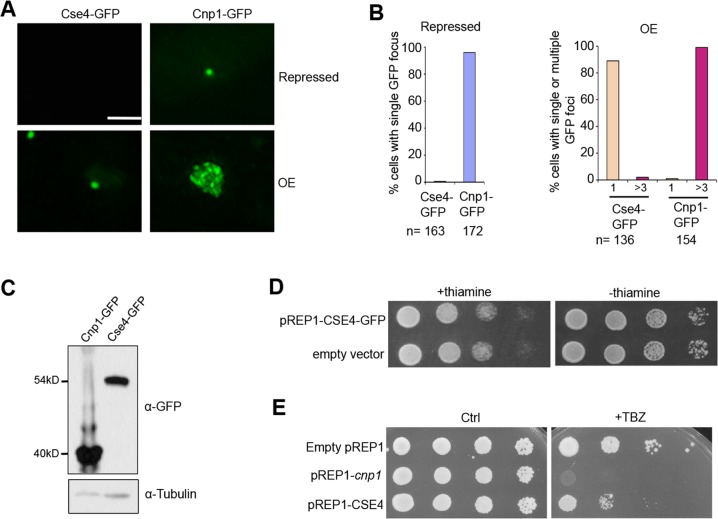
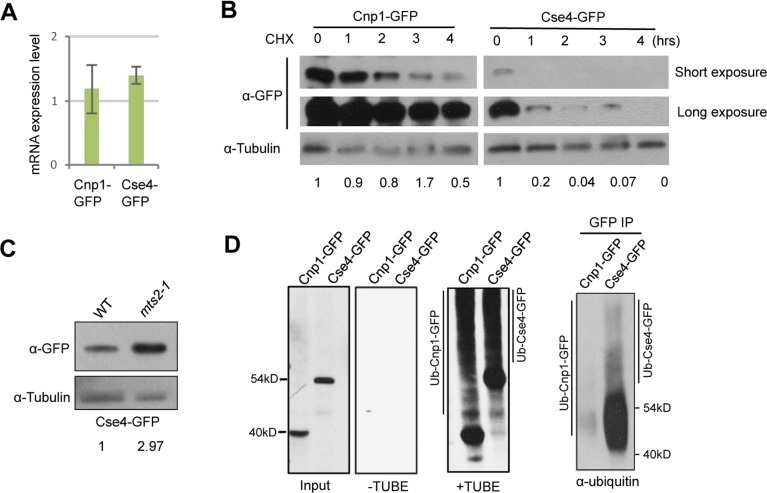
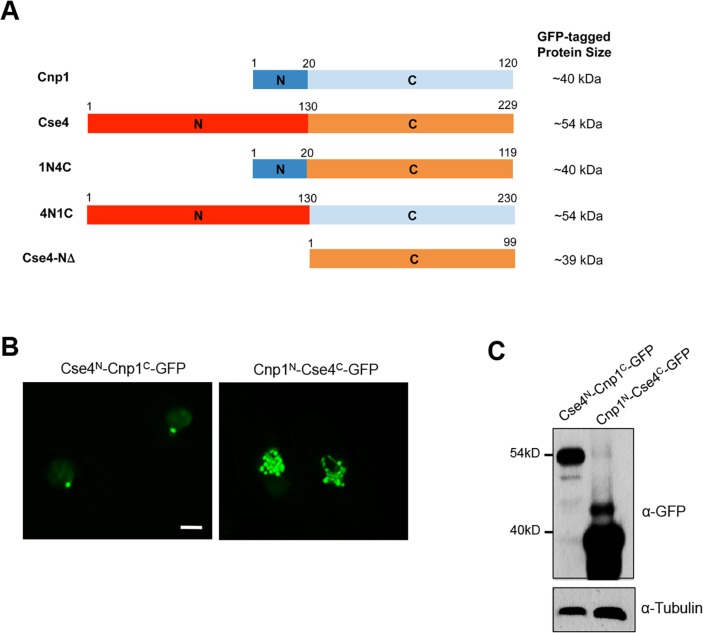
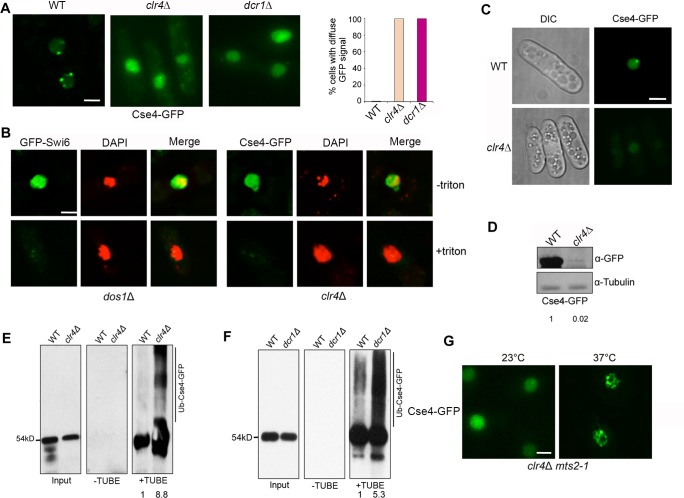
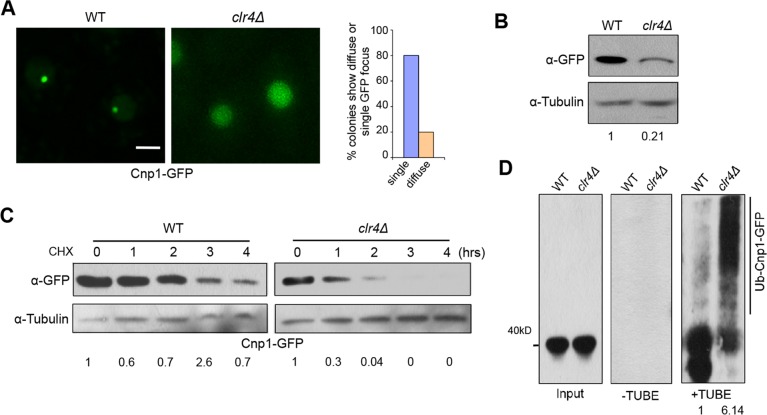
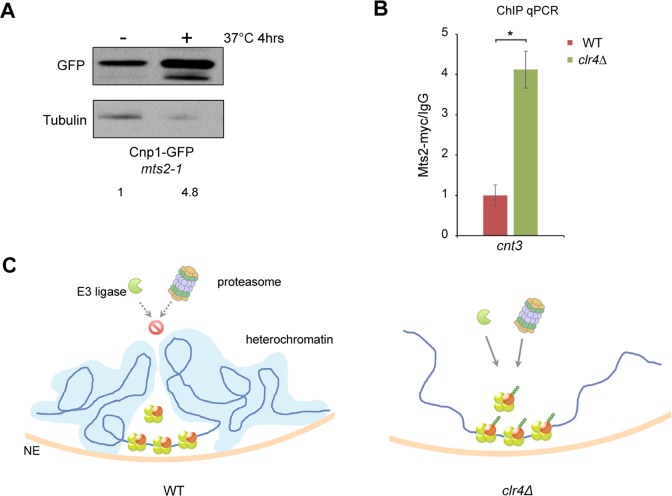
Centromere is a specialized chromatin domain that plays a vital role in chromosome segregation. In most eukaryotes, centromere is surrounded by the epigenetically distinct heterochromatin domain. Heterochromatin has been shown to contribute to centromere function, but the precise role of heterochromatin in centromere specification remains elusive. Centromeres in most eukaryotes, including fission yeast (Schizosaccharomyces pombe), are defined epigenetically by the histone H3 (H3) variant CENP-A. In contrast, the budding yeast Saccharomyces cerevisiae has genetically-defined point centromeres. The transition between regional centromeres and point centromeres is considered as one of the most dramatic evolutionary events in centromere evolution. Here we demonstrated that Cse4, the budding yeast CENP-A homolog, can localize to centromeres in fission yeast and partially substitute fission yeast CENP-ACnp1. But overexpression of Cse4 results in its localization to heterochromatic regions. Cse4 is subject to efficient ubiquitin-dependent degradation in S. pombe, and its N-terminal domain dictates its centromere distribution via ubiquitination. Notably, without heterochromatin and RNA interference (RNAi), Cse4 fails to associate with centromeres. We showed that RNAi-dependent heterochromatin mediates centromeric localization of Cse4 by protecting Cse4 from ubiquitin-dependent degradation. Heterochromatin also contributes to the association of native CENP-ACnp1 with centromeres via the same mechanism. These findings suggest that protection of CENP-A from degradation by heterochromatin is a general mechanism used for centromere assembly, and also provide novel insights into centromere evolution.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Histone H4 Facilitates the Proteolysis of the Budding Yeast CENP-ACse4 Centromeric Histone Variant.Genetics. 2017 Jan;205(1):113-124. doi: 10.1534/genetics.116.194027. Epub 2016 Oct 28. Genetics. 2017. PMID: 27794026 Free PMC article.
-
A Genome-Wide Screen Reveals a Role for the HIR Histone Chaperone Complex in Preventing Mislocalization of Budding Yeast CENP-A.Genetics. 2018 Sep;210(1):203-218. doi: 10.1534/genetics.118.301305. Epub 2018 Jul 16. Genetics. 2018. PMID: 30012561 Free PMC article.
-
The Ino80 complex mediates epigenetic centromere propagation via active removal of histone H3.Nat Commun. 2017 Sep 13;8(1):529. doi: 10.1038/s41467-017-00704-3. Nat Commun. 2017. PMID: 28904333 Free PMC article.
-
Protein kinases in mitotic phosphorylation of budding yeast CENP-A.Curr Genet. 2019 Dec;65(6):1325-1332. doi: 10.1007/s00294-019-00997-5. Epub 2019 May 22. Curr Genet. 2019. PMID: 31119371 Review.
-
Heterochromatin tells CENP-A where to go.Bioessays. 2008 Jun;30(6):526-9. doi: 10.1002/bies.20763. Bioessays. 2008. PMID: 18478529 Review.
Cited by
-
Repetitive Elements Contribute to the Diversity and Evolution of Centromeres in the Fungal Genus Verticillium.mBio. 2020 Sep 8;11(5):e01714-20. doi: 10.1128/mBio.01714-20. mBio. 2020. PMID: 32900804 Free PMC article.
-
Stable inheritance of CENP-A chromatin: Inner strength versus dynamic control.J Cell Biol. 2020 Oct 5;219(10):e202005099. doi: 10.1083/jcb.202005099. J Cell Biol. 2020. PMID: 32931551 Free PMC article.
-
Cell cycle control of kinetochore assembly.Nucleus. 2022 Dec;13(1):208-220. doi: 10.1080/19491034.2022.2115246. Nucleus. 2022. PMID: 36037227 Free PMC article. Review.
-
Centromeric and ectopic assembly of CENP-A chromatin in health and cancer: old marks and new tracks.Nucleic Acids Res. 2019 Feb 20;47(3):1051-1069. doi: 10.1093/nar/gky1298. Nucleic Acids Res. 2019. PMID: 30590707 Free PMC article. Review.
-
Rbm10 facilitates heterochromatin assembly via the Clr6 HDAC complex.Epigenetics Chromatin. 2021 Jan 19;14(1):8. doi: 10.1186/s13072-021-00382-y. Epigenetics Chromatin. 2021. PMID: 33468217 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
